

Convergent Epigenetic Mechanisms Avoid Constitutive Expression of Immune Receptor Gene Subsets
- PMID: 34557212
- PMCID: PMC8452986
- DOI: 10.3389/fpls.2021.703667
Convergent Epigenetic Mechanisms Avoid Constitutive Expression of Immune Receptor Gene Subsets
Abstract
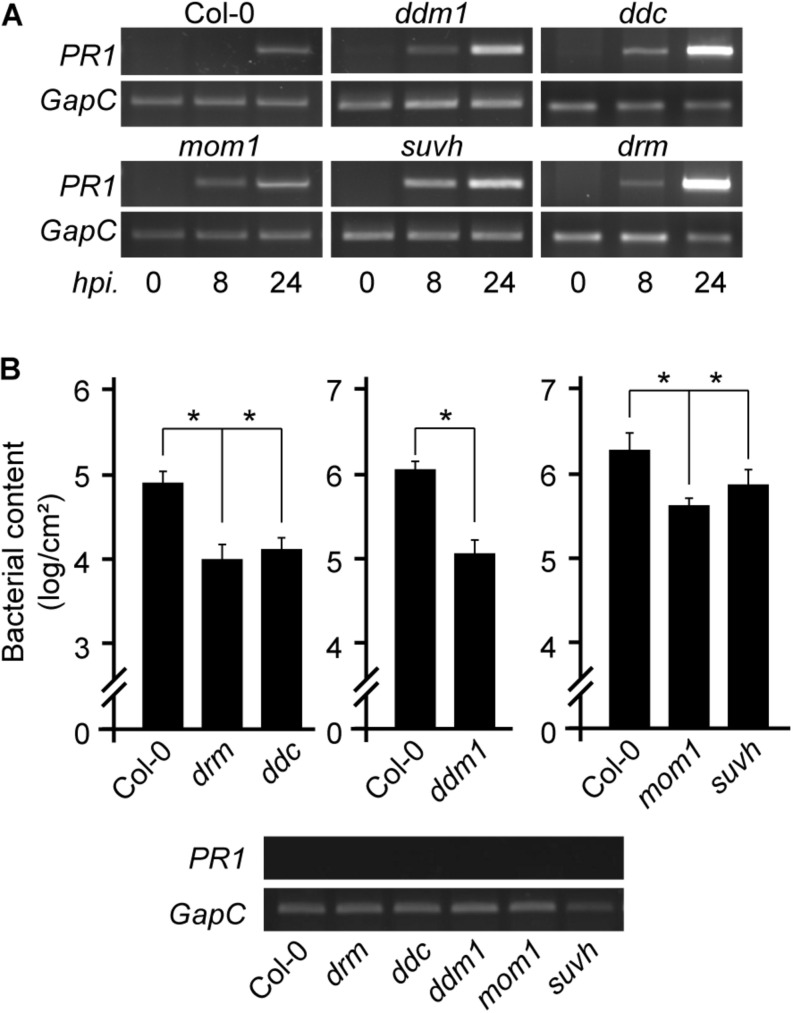
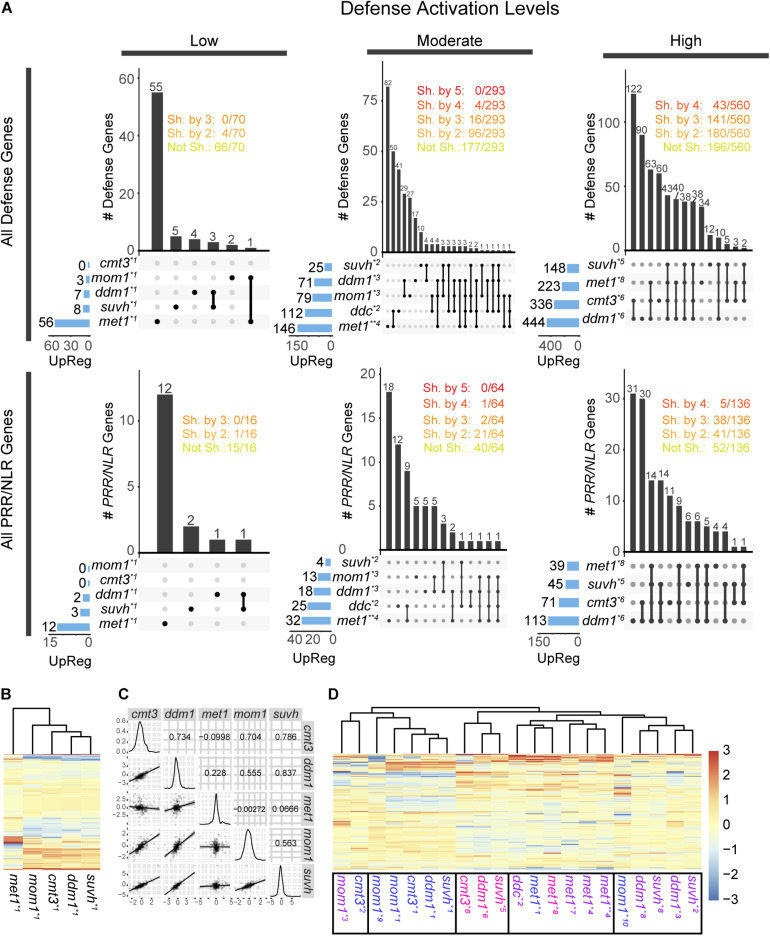
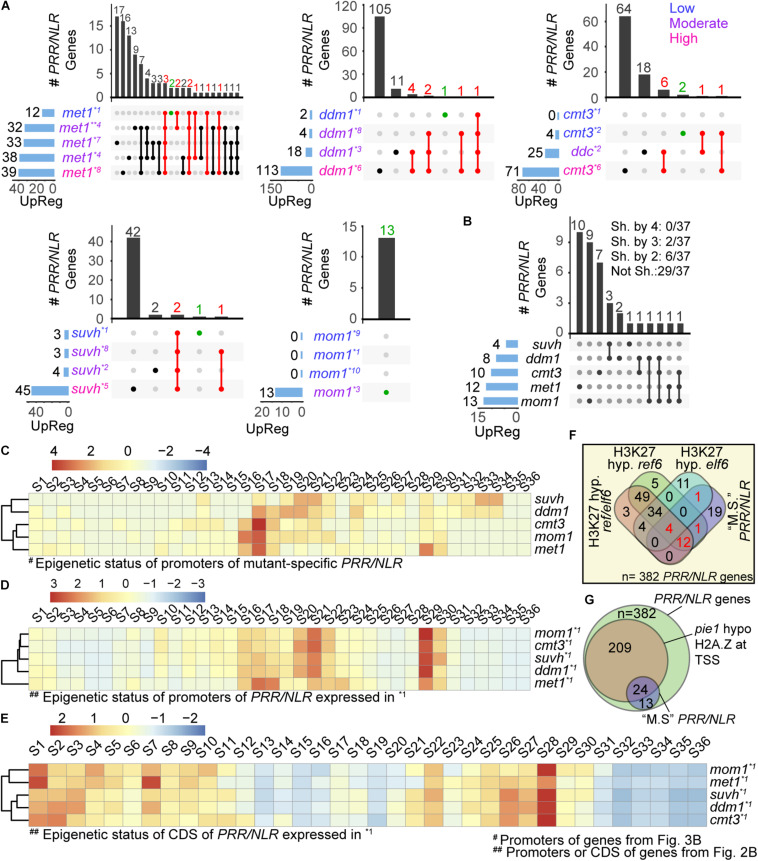
The gene pool encoding PRR and NLR immune receptors determines the ability of a plant to resist microbial infections. Basal expression of these genes is prevented by diverse mechanisms since their hyperactivity can be harmful. To approach the study of epigenetic control of PRR/NLR genes we here analyzed their expression in mutants carrying abnormal repressive 5-methyl cytosine (5-mC) and histone 3 lysine 9 dimethylation (H3K9me2) marks, due to lack of MET1, CMT3, MOM1, SUVH4/5/6, or DDM1. At optimal growth conditions, none of the mutants showed basal expression of the defense gene marker PR1, but all of them had greater resistance to Pseudomonas syringae pv. tomato than wild type plants, suggesting they are primed to stimulate immune cascades. Consistently, analysis of available transcriptomes indicated that all mutants showed activation of particular PRR/NLR genes under some growth conditions. Under low defense activation, 37 PRR/NLR genes were expressed in these plants, but 29 of them were exclusively activated in specific mutants, indicating that MET1, CMT3, MOM1, SUVH4/5/6, and DDM1 mediate basal repression of different subsets of genes. Some epigenetic marks present at promoters, but not gene bodies, could explain the activation of these genes in the mutants. As expected, suvh4/5/6 and ddm1 activated genes carrying 5-mC and H3K9me2 marks in wild type plants. Surprisingly, all mutants expressed genes harboring promoter H2A.Z/H3K27me3 marks likely affected by the chromatin remodeler PIE1 and the histone demethylase REF6, respectively. Therefore, MET1, CMT3, MOM1, SUVH4/5/6, and DDM1, together with REF6, seemingly contribute to the establishment of chromatin states that prevent constitutive PRR/NLR gene activation, but facilitate their priming by modulating epigenetic marks at their promoters.
Keywords: 5-mC/H3K9me2 and H2A.Z/H3K27me3 marks; PRR/NLR immune receptor genes; defense cascades; epigenetics; priming.
Copyright © 2021 Cambiagno, Torres and Alvarez.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
LinkOut - more resources
Full Text Sources
Research Materials