

Mechanisms of Plant Responses and Adaptation to Soil Salinity
- PMID: 34557705
- PMCID: PMC8454569
- DOI: 10.1016/j.xinn.2020.100017
Mechanisms of Plant Responses and Adaptation to Soil Salinity
Abstract
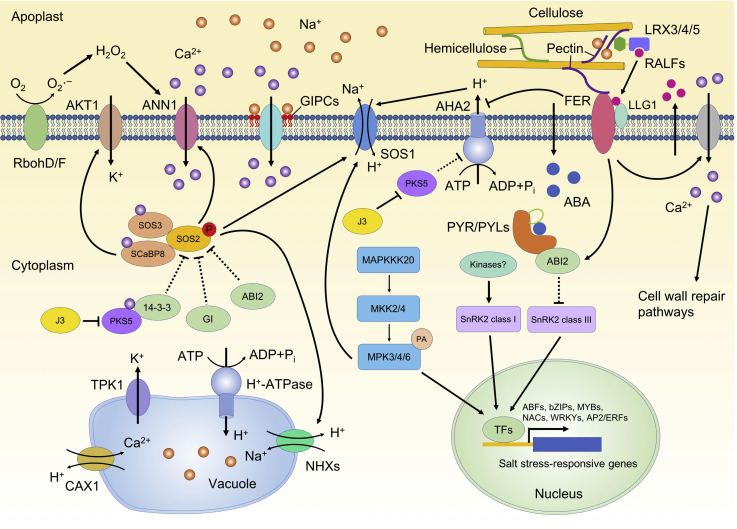
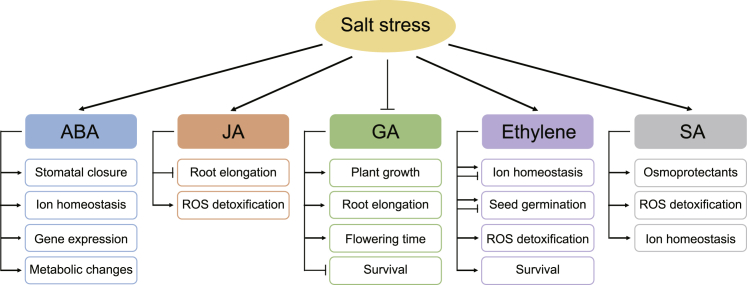
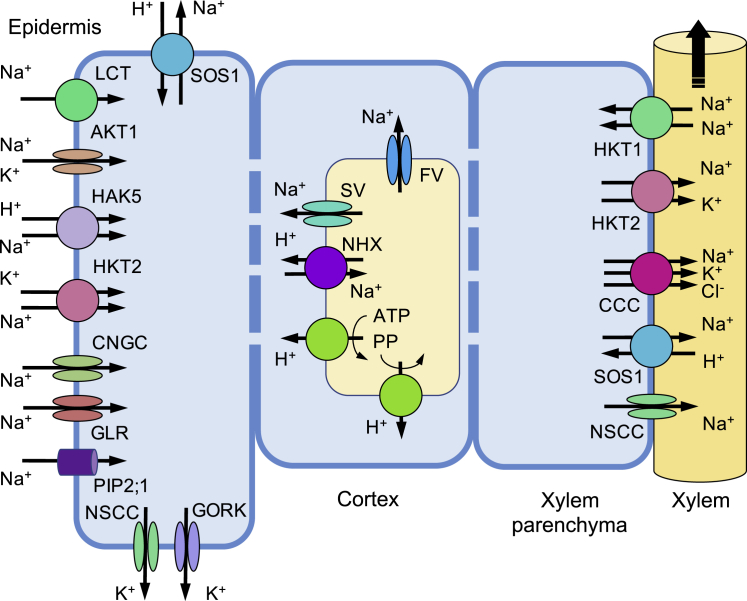
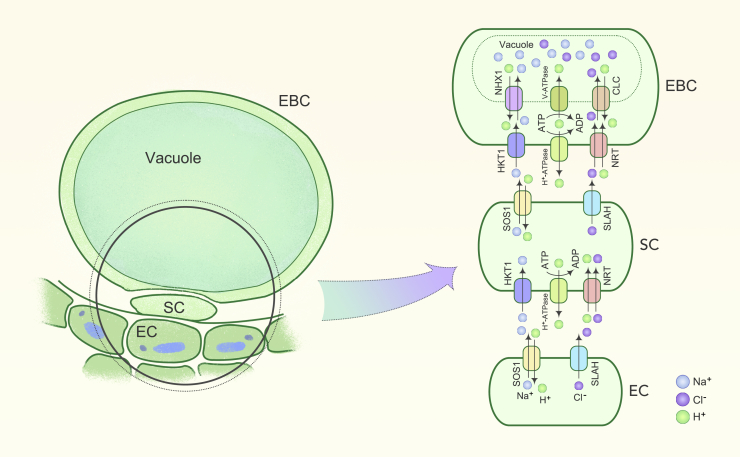
Soil salinity is a major environmental stress that restricts the growth and yield of crops. Understanding the physiological, metabolic, and biochemical responses of plants to salt stress and mining the salt tolerance-associated genetic resource in nature will be extremely important for us to cultivate salt-tolerant crops. In this review, we provide a comprehensive summary of the mechanisms of salt stress responses in plants, including salt stress-triggered physiological responses, oxidative stress, salt stress sensing and signaling pathways, organellar stress, ion homeostasis, hormonal and gene expression regulation, metabolic changes, as well as salt tolerance mechanisms in halophytes. Important questions regarding salt tolerance that need to be addressed in the future are discussed.
Keywords: halophyte; hormones; ion homeostasis; osmotic stress; oxidative stress; salt stress; salt stress sensing.
© 2020.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- FAO . Food and Agriculture Organization of the United Nations and Earthscan; 2011. The State of the World’s Land and Water Resources for Food and Agriculture (SOLAW)—Managing Systems at Risk.http://www.fao.org/3/a-i1688e.pdf
-
- Rengasamy P. World salinization with emphasis on Australia. J. Exp. Bot. 2006;57:1017–1023. - PubMed
-
- Munns R., Tester M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008;59:651–681. - PubMed
Publication types
LinkOut - more resources
Full Text Sources
