

The Arabidopsis thaliana LysM-containing Receptor-Like Kinase 2 is required for elicitor-induced resistance to pathogens
- PMID: 34558681
- PMCID: PMC9293440
- DOI: 10.1111/pce.14192
The Arabidopsis thaliana LysM-containing Receptor-Like Kinase 2 is required for elicitor-induced resistance to pathogens
Abstract
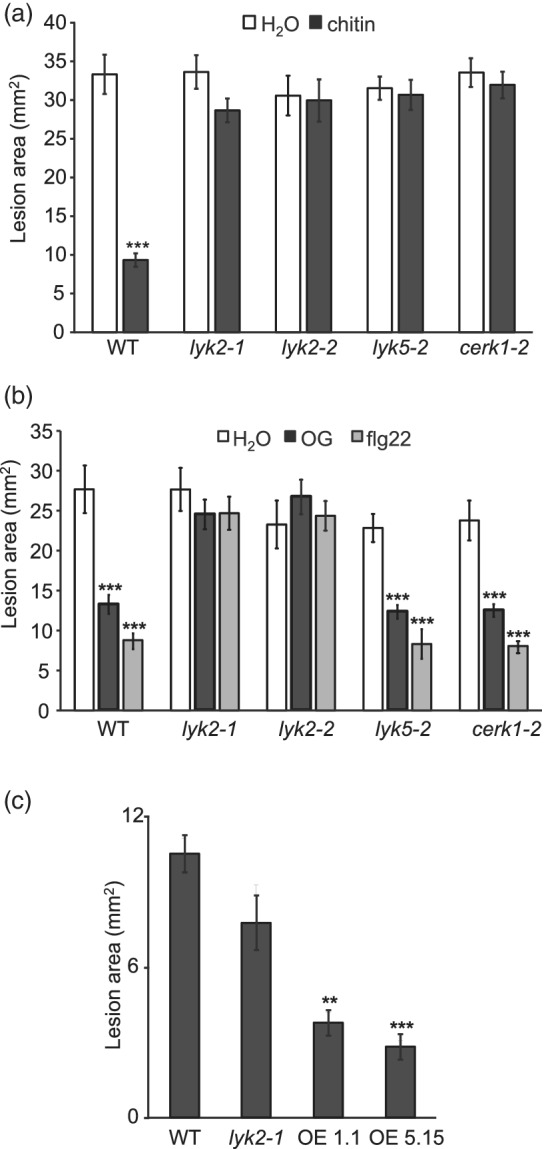
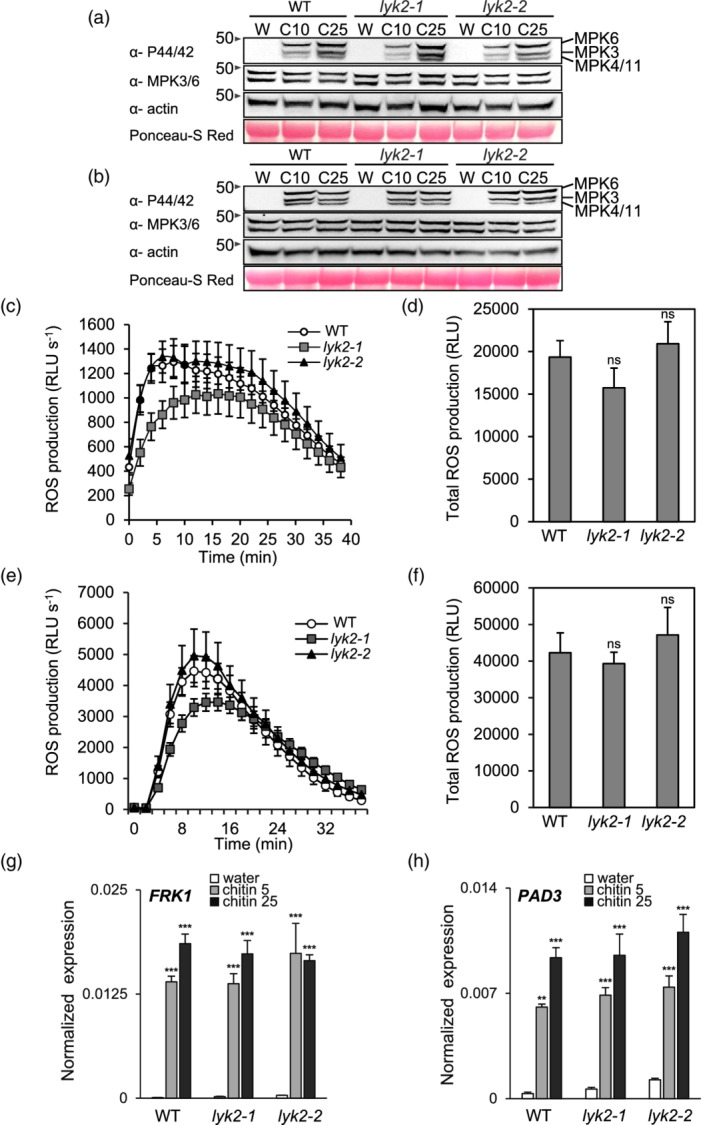
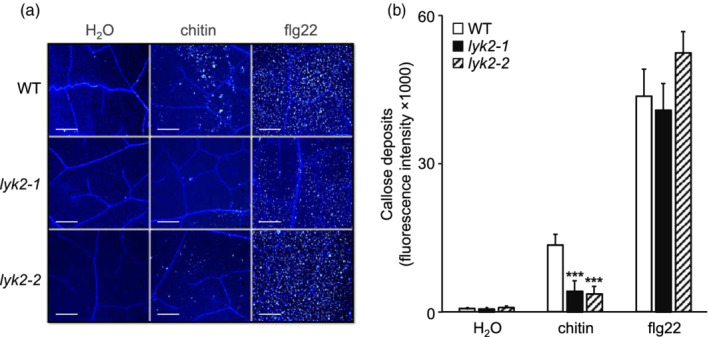
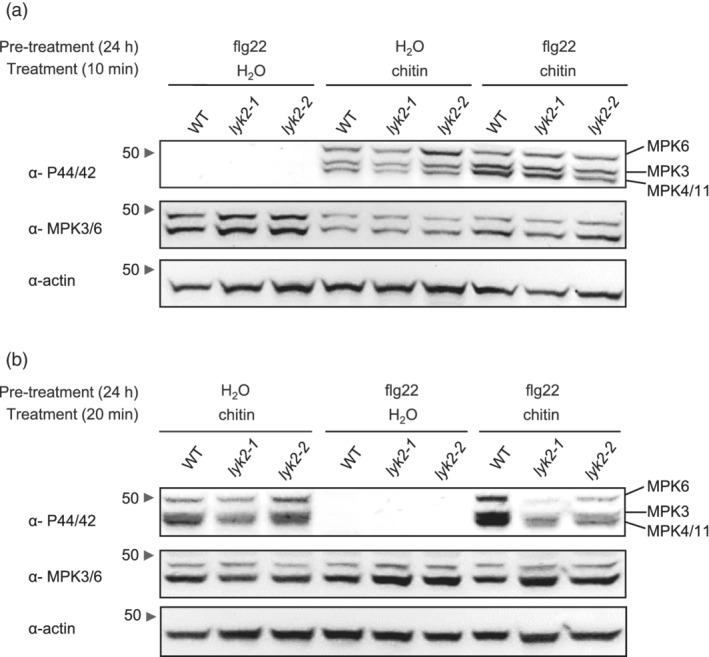
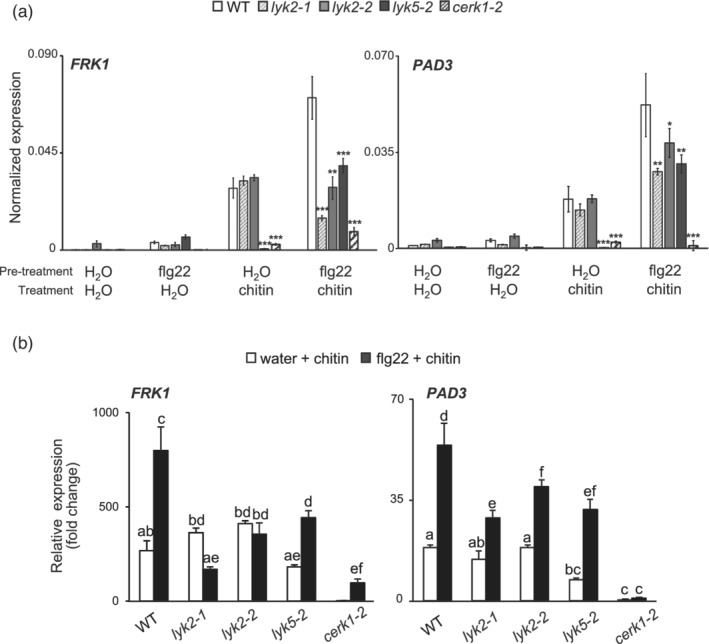
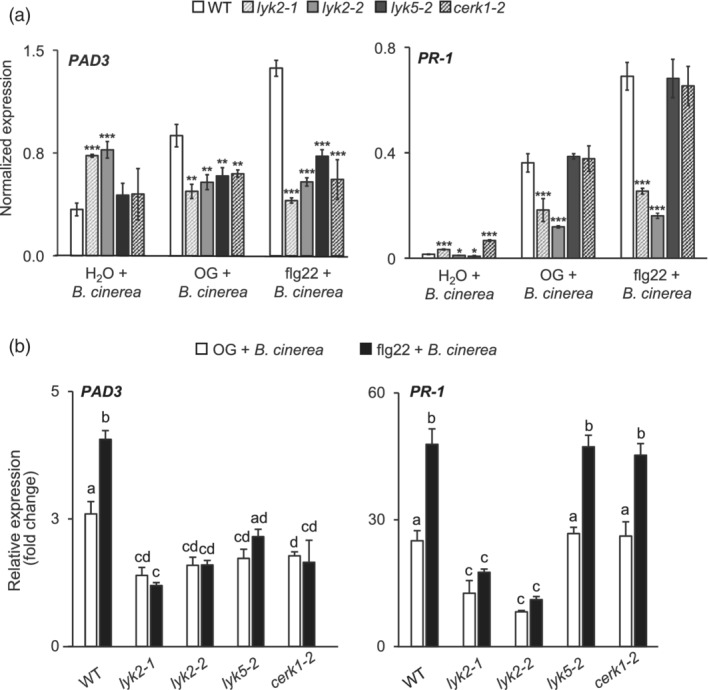
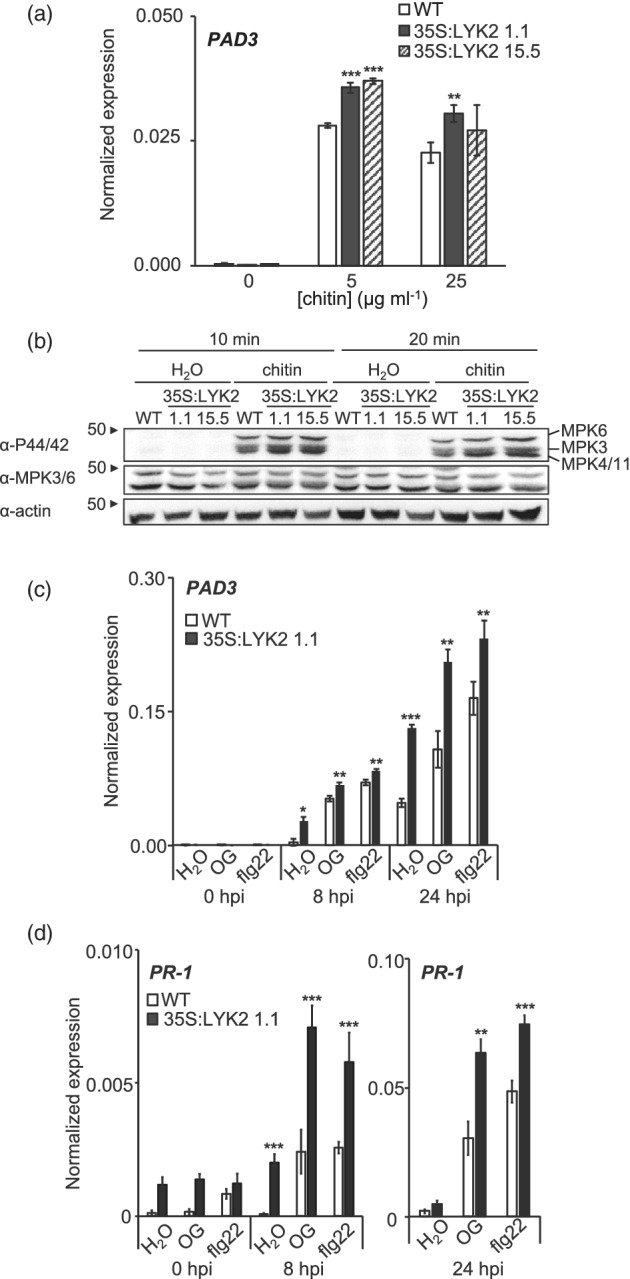
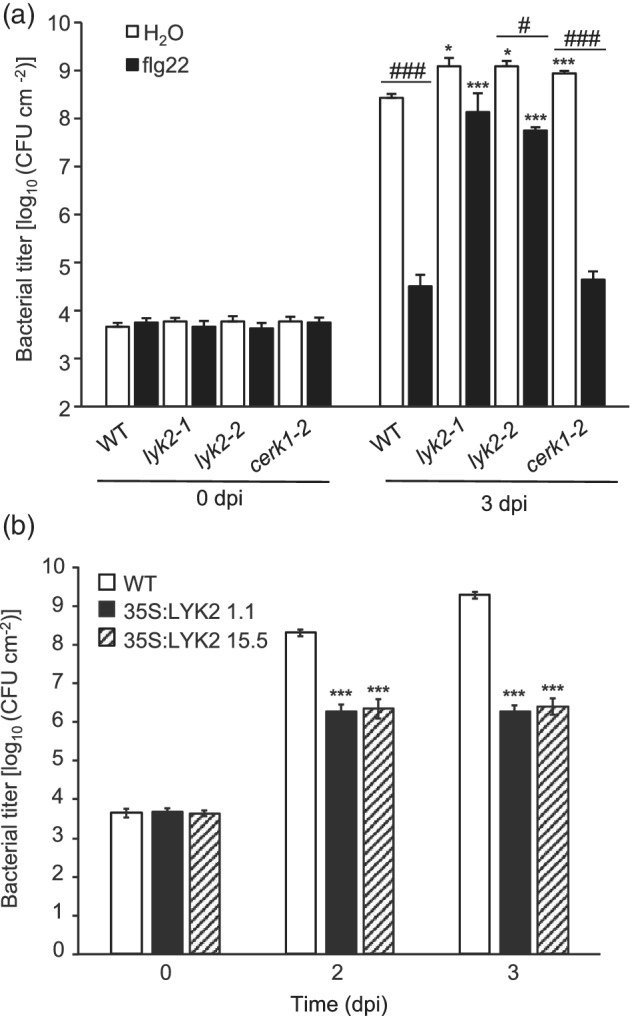
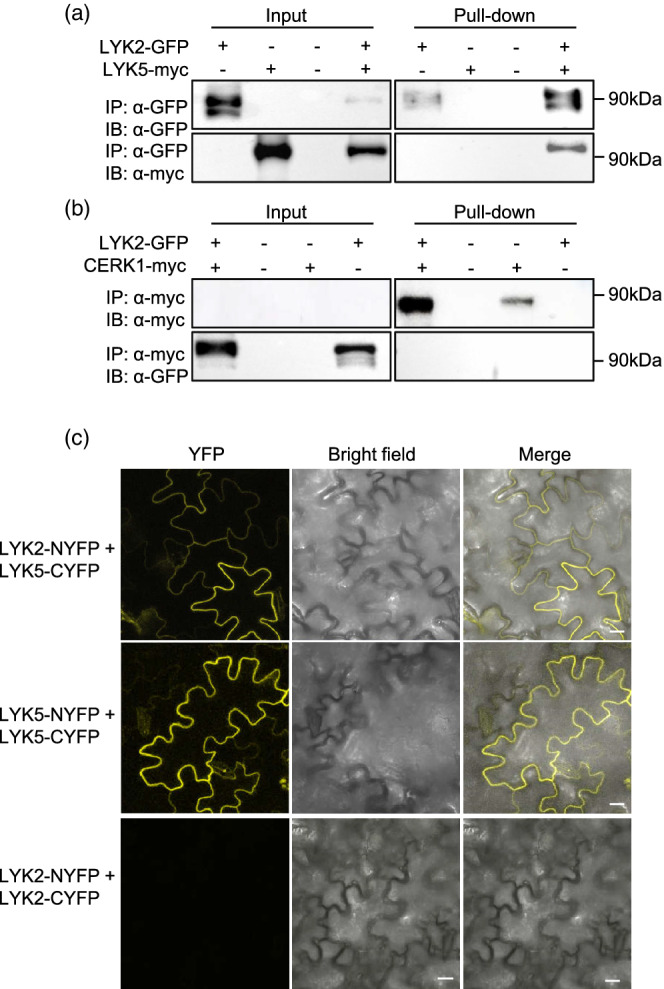
In Arabidopsis thaliana, perception of chitin from fungal cell walls is mediated by three LysM-containing Receptor-Like Kinases (LYKs): CERK1, which is absolutely required for chitin perception, and LYK4 and LYK5, which act redundantly. The role in plant innate immunity of a fourth LYK protein, LYK2, is currently not known. Here we show that CERK1, LYK2 and LYK5 are dispensable for basal susceptibility to B. cinerea but are necessary for chitin-induced resistance to this pathogen. LYK2 is dispensable for chitin perception and early signalling events, though it contributes to callose deposition induced by this elicitor. Notably, LYK2 is also necessary for enhanced resistance to B. cinerea and Pseudomonas syringae induced by flagellin and for elicitor-induced priming of defence gene expression during fungal infection. Consistently, overexpression of LYK2 enhances resistance to B. cinerea and P. syringae and results in increased expression of defence-related genes during fungal infection. LYK2 appears to be required to establish a primed state in plants exposed to biotic elicitors, ensuring a robust resistance to subsequent pathogen infections.
Keywords: Botrytis cinerea; chitin; plant innate immunity; priming.
© 2021 The Authors. Plant, Cell & Environment published by John Wiley & Sons Ltd.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures
References
-
- An, H. J. , Lurie, S. , Greve, L. C. , Rosenquist, D. , Kirmiz, C. , Labavitch, J. M. , & Lebrilla, C. B. (2005). Determination of pathogen‐related enzyme action by mass spectrometry analysis of pectin breakdown products of plant cell walls. Analytical Biochemistry, 338(1), 71–82. 10.1016/j.ab.2004.11.004 - DOI - PubMed
-
- Arrighi, J. , Barre, A. , Ben Amor, B. , Bersoult, A. , Soriano, L. , Mirabella, R. , … Gough, C. (2006). The Medicago truncatula lysin [corrected] motif‐receptor‐like kinase gene family includes NFP and new nodule‐expressed genes. Plant Physiology, 142(1), 265–279. 10.1104/pp.106.084657 - DOI - PMC - PubMed
-
- Asai, T. , Tena, G. , Plotnikova, J. , Willmann, M. R. , Chiu, W. L. , Gomez‐Gomez, L. , … Sheen, J. (2002). MAP kinase signalling cascade in Arabidopsis innate immunity. Nature, 415(6875), 977–983. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
