

UFMylation of MRE11 is essential for telomere length maintenance and hematopoietic stem cell survival
- PMID: 34559557
- PMCID: PMC8462904
- DOI: 10.1126/sciadv.abc7371
UFMylation of MRE11 is essential for telomere length maintenance and hematopoietic stem cell survival
Abstract
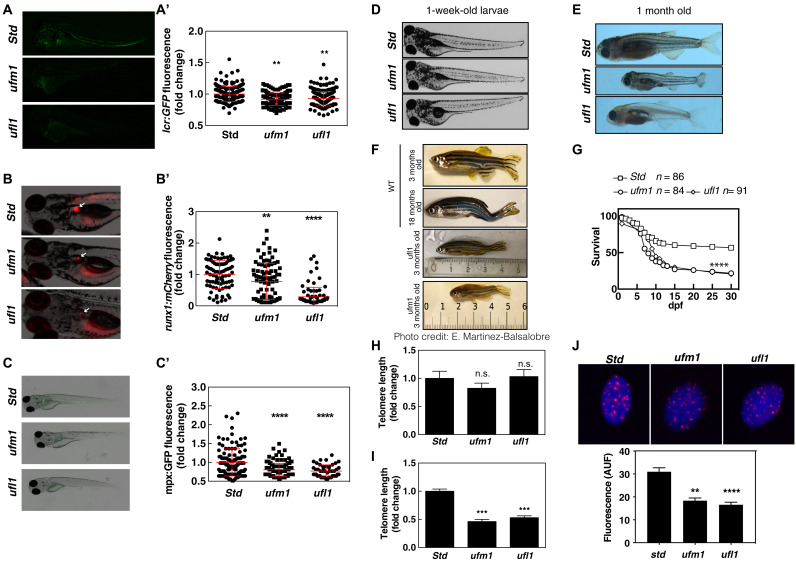
Ubiquitin-fold modifier 1 (UFM1) is involved in neural and erythroid development, yet its biological roles in these processes are unknown. Here, we generated zebrafish models deficient in Ufm1 and Ufl1 that exhibited telomere shortening associated with developmental delay, impaired hematopoiesis and premature aging. We further report that HeLa cells lacking UFL1 have instability of telomeres replicated by leading-strand synthesis. We uncover that MRE11 UFMylation is necessary for the recruitment of the phosphatase PP1-α leading to dephosphorylation of NBS1. In the absence of UFMylation, NBS1 remains phosphorylated, thereby reducing MRN recruitment to telomeres. The absence of MRN at telomeres favors the formation of the TRF2-Apollo/SNM1 complex consistent with the loss of leading telomeres. These results suggest that MRE11-UFMylation may serve as module to recruit PP1-α. Last, zebrafish expressing Mre11 that cannot be UFMylated phenocopy Ufm1-deficient zebrafish, demonstrating that UFMylation of MRE11 is a previously undescribed evolutionarily conserved mechanisms regulating telomere length.
Figures





References
-
- Zhang M., Zhu X., Zhang Y., Cai Y., Chen J., Sivaprakasam S., Gurav A., Pi W., Makala L., Wu J., Pace B., Tuan-Lo D., Ganapathy V., Singh N., Li H., RCAD/Ufl1, a Ufm1 E3 ligase, is essential for hematopoietic stem cell function and murine hematopoiesis. Cell Death Differ. 22, 1922–1934 (2015). - PMC - PubMed
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
