

Discovery and functional characterization of N-(thiazol-2-yl)-benzamide analogs as the first class of selective antagonists of the Zinc-Activated Channel (ZAC)
- PMID: 34560054
- PMCID: PMC9979163
- DOI: 10.1016/j.bcp.2021.114782
Discovery and functional characterization of N-(thiazol-2-yl)-benzamide analogs as the first class of selective antagonists of the Zinc-Activated Channel (ZAC)
Abstract
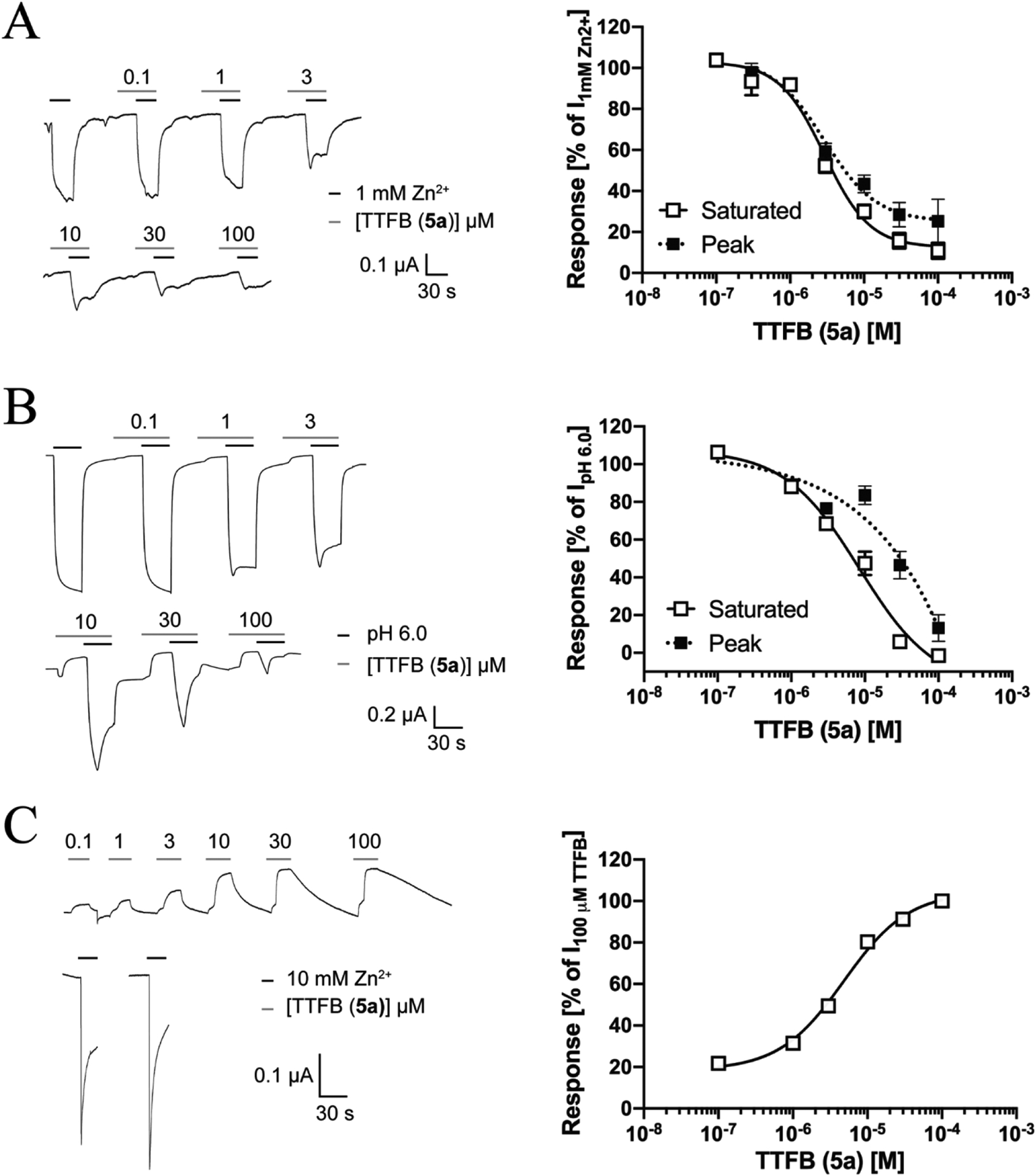
The Zinc-Activated Channel (ZAC) is an atypical member of the Cys-loop receptor (CLR) superfamily of pentameric ligand-gated ion channels, with its very different endogenous agonists and signalling properties. In this study, a compound library screening at ZAC resulted in the identification of 2-(5-bromo-2-chlorobenzamido)-4-methylthiazole-5-methyl ester (1) as a novel ZAC antagonist. The structural determinants for ZAC activity in 1 were investigated by functional characterization of 61 analogs at ZAC expressed in Xenopus oocytes by two-electrode voltage clamp electrophysiology, and couple of analogs exerting more potent ZAC inhibition than 1 were identified (IC50 values: 1-3 μM). 1 and N-(4-(tert-butyl)thiazol-2-yl)-3-fluorobenzamide (5a, TTFB) were next applied in studies of the functional properties and the mode of action of this novel class of ZAC antagonists. TTFB was a roughly equipotent antagonist of Zn+- and H+-evoked ZAC signaling and of spontaneous ZAC activity, and the slow on-set of its channel block suggested that its ZAC inhibition is state-dependent. TTFB was found to be a selective ZAC antagonist, exhibiting no significant agonist, antagonist or modulatory activity at 5-HT3A, α3β4 nicotinic acetylcholine, α1β2γ2S GABAA or α1 glycine receptors at 30 μM. 1 displayed largely non-competitive antagonism of Zn2+-induced ZAC signalling, and TTFB was demonstrated to target the transmembrane and/or intracellular domains of the receptor, which collectively suggests that the N-(thiazol-2-yl)-benzamide analog acts a negative allosteric modulator of ZAC. We propose that this first class of selective ZAC antagonists could constitute useful pharmacological tools in future explorations of the presently poorly elucidated physiological functions governed by this CLR.
Keywords: Cys-loop receptor (CLR); N-(thiazol-2-yl)-benzamide analogs; Negative allosteric modulator (NAM); Pentameric ligand-gated ion channel (pLGIC); State-dependent inhibition; Zinc-Activated Channel (ZAC).
Copyright © 2021 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures









References
-
- Walstab J, Rappold G, Niesler B, 5-HT3 receptors: role in disease and target of drugs, Pharmacol Ther 128 (1) (2010) 146–169. - PubMed
-
- Fakhfouri G, Rahimian R, Dyhrfjeld-Johnsen J, Zirak MR, Beaulieu J-M, Witkin JM, 5-HT3 receptor antagonists in neurologic and neuropsychiatric disorders: The iceberg still lies beneath the surface, Pharmacol Rev 71 (3) (2019) 383–412. - PubMed
-
- Lynch JW, Zhang Y, Talwar S, Estrada-Mondragon A, Glycine Receptor Drug Discovery, Adv Pharmacol 79 (2017) 225–253. - PubMed
-
- Chua HC, Chebib M, GABAA Receptors and the Diversity in their Structure and Pharmacology, Adv Pharmacol 79 (2017) 1–34. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
