

An atomistic model of the coronavirus replication-transcription complex as a hexamer assembled around nsp15
- PMID: 34562452
- PMCID: PMC8494237
- DOI: 10.1016/j.jbc.2021.101218
An atomistic model of the coronavirus replication-transcription complex as a hexamer assembled around nsp15
Abstract
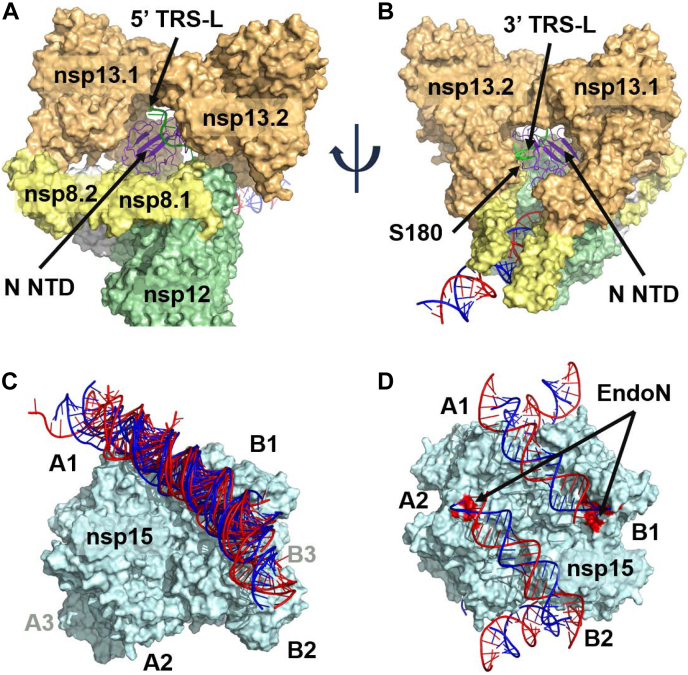
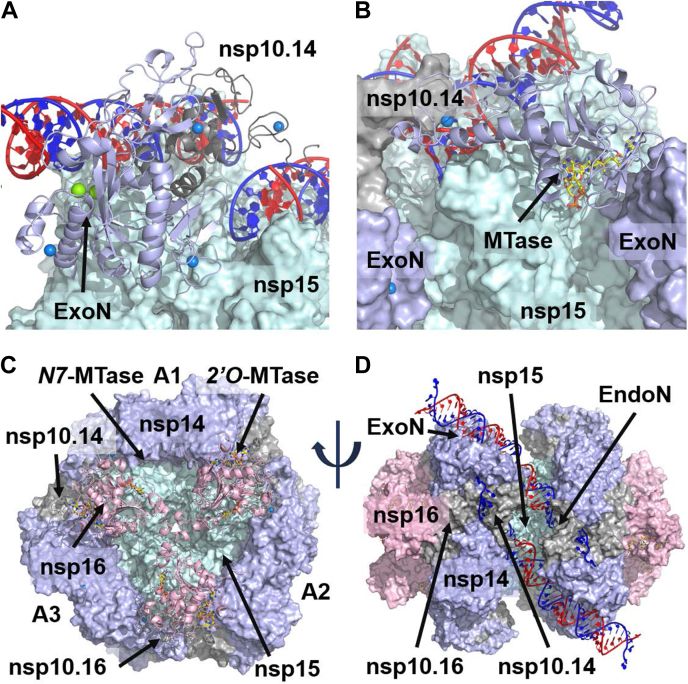
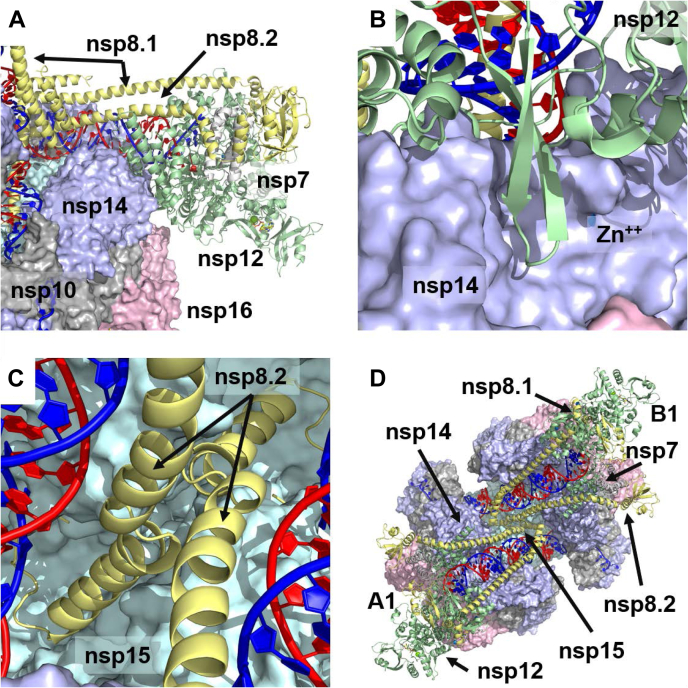
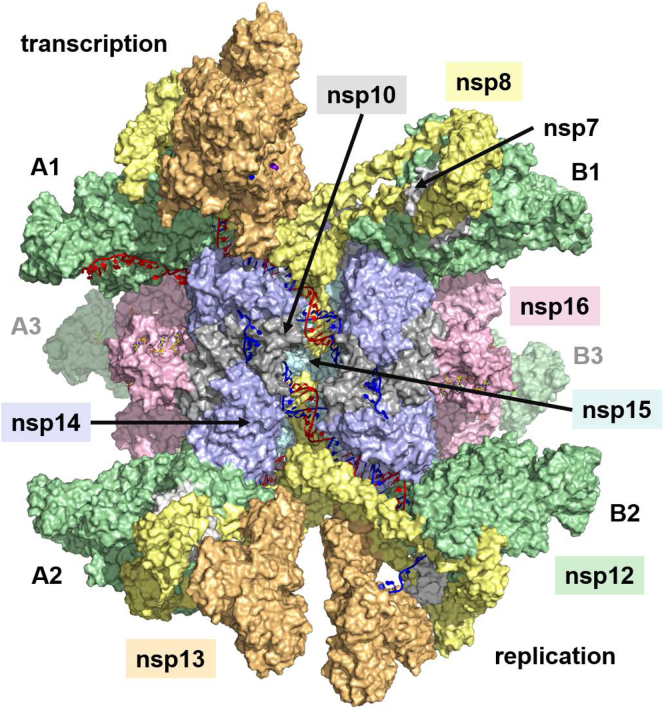
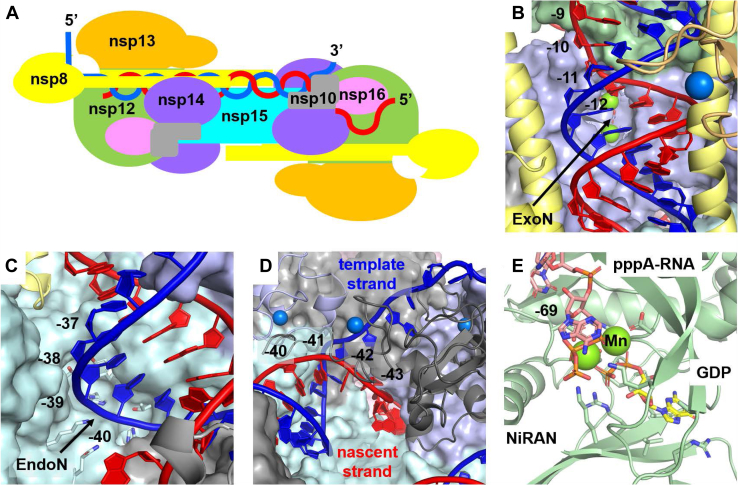
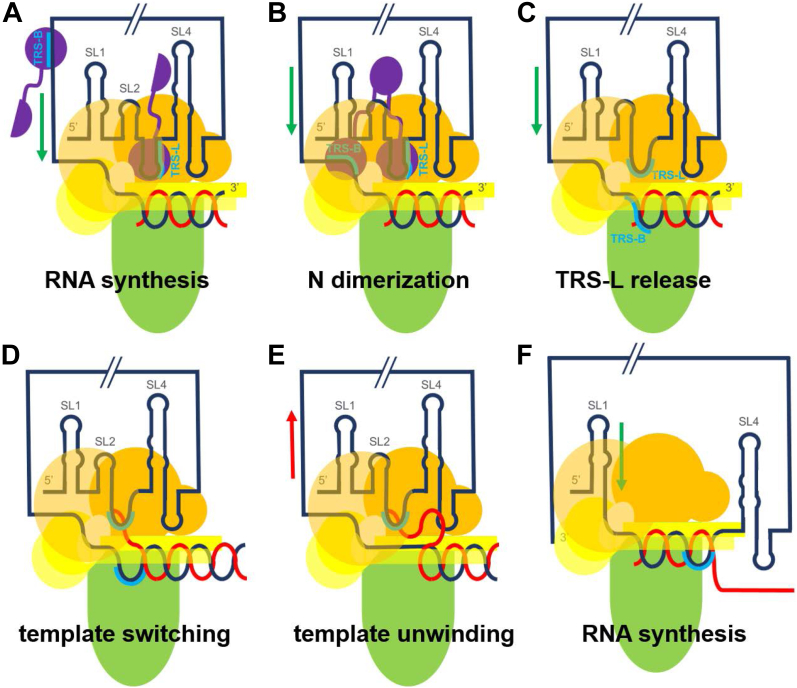
The SARS-CoV-2 replication-transcription complex is an assembly of nonstructural viral proteins that collectively act to reproduce the viral genome and generate mRNA transcripts. While the structures of the individual proteins involved are known, how they assemble into a functioning superstructure is not. Applying molecular modeling tools, including protein-protein docking, to the available structures of nsp7-nsp16 and the nucleocapsid, we have constructed an atomistic model of how these proteins associate. Our principal finding is that the complex is hexameric, centered on nsp15. The nsp15 hexamer is capped on two faces by trimers of nsp14/nsp16/(nsp10)2, which then recruit six nsp12/nsp7/(nsp8)2 polymerase subunits to the complex. To this, six subunits of nsp13 are arranged around the superstructure, but not evenly distributed. Polymerase subunits that coordinate dimers of nsp13 are capable of binding the nucleocapsid, which positions the 5'-UTR TRS-L RNA over the polymerase active site, a state distinguishing transcription from replication. Analysis of the viral RNA path through the complex indicates the dsRNA that exits the polymerase passes over the nsp14 exonuclease and nsp15 endonuclease sites before being unwound by a convergence of zinc fingers from nsp10 and nsp14. The template strand is then directed away from the complex, while the nascent strand is directed to the sites responsible for mRNA capping. The model presents a cohesive picture of the multiple functions of the coronavirus replication-transcription complex and addresses fundamental questions related to proofreading, template switching, mRNA capping, and the role of the endonuclease.
Keywords: SARS-CoV-2; coronavirus; molecular modeling; nsp15; proofreading; protein structure; structure model; viral replication; viral transcription.
Copyright © 2021 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures
References
-
- Hillen H.S., Kokic G., Farnung L., Dienemann C., Tegunov D., Cramer P. Structure of replicating SARS-CoV-2 polymerase. Nature. 2020;584:154–156. - PubMed
-
- Lehmann K.C., Gulyaeva A., Zevenhoven-Dobbe J.C., Janssen G.M., Ruben M., Overkleeft H.S., van Veelen P.A., Samborskiy D.V., Kravchenko A.A., Leontovich A.M., Sidorov I.A., Snijder E.J., Posthuma C.C., Gorbalenya A.E. Discovery of an essential nucleotidylating activity associated with a newly delineated conserved domain in the RNA polymerase-containing protein of all nidoviruses. Nucleic Acids Res. 2015;43:8416–8434. - PMC - PubMed
-
- Slanina H., Madhugiri R., Bylapudi G., Schultheiss K., Karl N., Gulyaeva A., Gorbalenya A.E., Linne U., Ziebuhr J. Coronavirus replication-transcription complex: Vital and selective NMPylation of a conserved site in nsp9 by the NiRAN-RdRp subunit. Proc. Natl. Acad. Sci. U. S. A. 2021;118 - PMC - PubMed
-
- Yan L., Ge J., Zheng L., Zhang Y., Gao Y., Wang T., Huang Y., Yang Y., Gao S., Li M., Liu Z., Wang H., Li Y., Chen Y., Guddat L.W. Cryo-EM structure of an extended SARS-CoV-2 replication and transcription complex reveals an intermediate state in cap synthesis. Cell. 2021;184:184–193.e110. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
