

Distance Dependent Contribution of Ants to Pollination but Not Defense in a Dioecious, Ambophilous Gymnosperm
- PMID: 34567036
- PMCID: PMC8459830
- DOI: 10.3389/fpls.2021.722405
Distance Dependent Contribution of Ants to Pollination but Not Defense in a Dioecious, Ambophilous Gymnosperm
Abstract
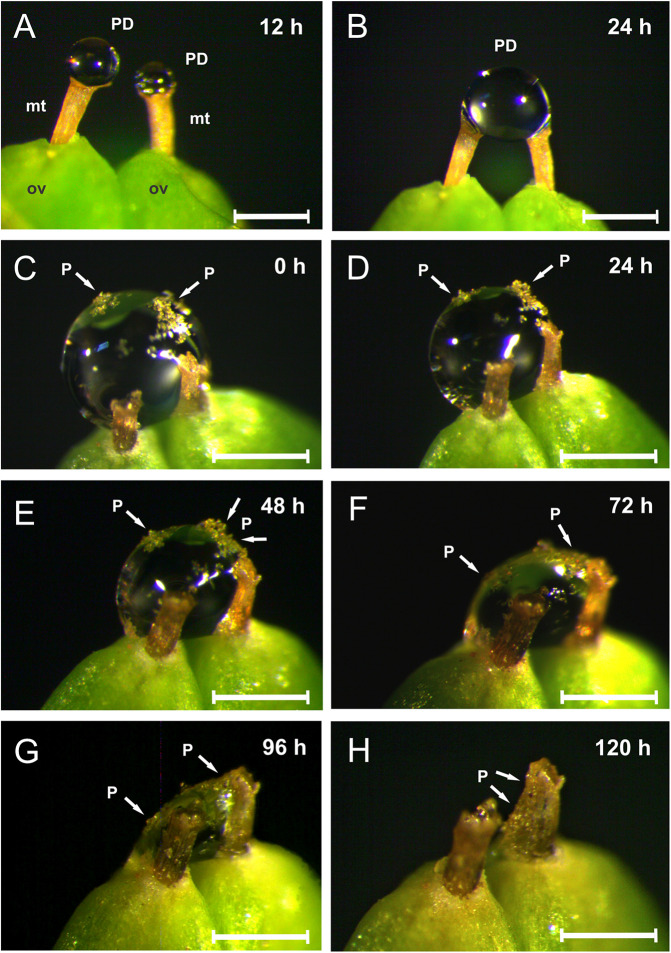
Dioecious plants are obligate outcrossers with separate male and female individuals, which can result in decreased seed set with increasing distance between the sexes. Wind pollination is a common correlate of dioecy, yet combined wind and insect pollination (ambophily) could be advantageous in compensating for decreased pollen flow to isolated females. Dioecious, ambophilous gymnosperms Ephedra (Gnetales) secrete pollination drops (PDs) in female cones that capture airborne pollen and attract ants that feed on them. Plant sugary secretions commonly reward ants in exchange for indirect plant defense against herbivores, and more rarely for pollination. We conducted field experiments to investigate whether ants are pollinators and/or plant defenders of South American Ephedra triandra, and whether their contribution to seed set and seed cone protection varies with distance between female and male plants. We quantified pollen flow in the wind and assessed the effectiveness of ants as pollinators by investigating their relative contribution to seed set, and their visitation rate in female plants at increasing distance from the nearest male. Ants accounted for most insect visits to female cones of E. triandra, where they consumed PDs, and pollen load was larger on bigger ants without reduction in pollen viability. While wind pollination was the main contributor to seed set overall, the relative contribution of ants was distance dependent. Ant contribution to seed set was not significant at shorter distances, yet at the farthest distance from the nearest male (23 m), where 20 times less pollen reached females, ants enhanced seed set by 30% compared to plants depending solely on wind pollination. We found no evidence that ants contribute to plant defense by preventing seed cone damage. Our results suggest that, despite their short-range movements, ants can offset pollen limitation in isolated females of wind-pollinated plants with separate sexes. We propose that ants enhance plant reproductive success via targeted delivery of airborne pollen, through frequent contact with ovule tips while consuming PDs. Our study constitutes the first experimental quantification of distance-dependent contribution of ants to pollination and provides a working hypothesis for ambophily in other dioecious plants lacking pollinator reward in male plants.
Keywords: Ephedra; Gnetales; ant pollination; dioecy; gymnosperm pollination; pollination drop; protective mutualism; wind pollination.
Copyright © 2021 Aranda-Rickert, Torréns, Yela, Brizuela and Di Stilio.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures





Similar articles
-
Influence of local density and sex ratio on pollination in an ambophilous flowering plant.Am J Bot. 2020 Apr;107(4):587-598. doi: 10.1002/ajb2.1453. Epub 2020 Mar 30. Am J Bot. 2020. PMID: 32227341
-
Phylogenetic and functional signals in gymnosperm ovular secretions.Ann Bot. 2017 Nov 28;120(6):923-936. doi: 10.1093/aob/mcx103. Ann Bot. 2017. PMID: 29045531 Free PMC article.
-
The best of two worlds: ecology and evolution of ambophilous plants.Biol Rev Camb Philos Soc. 2023 Apr;98(2):391-420. doi: 10.1111/brv.12911. Epub 2022 Oct 21. Biol Rev Camb Philos Soc. 2023. PMID: 36270973 Review.
-
Reproductive biology and pollination ecology of Triplaris gardneriana (Polygonaceae): a case of ambophily in the Brazilian Chaco.Plant Biol (Stuttg). 2017 Jul;19(4):504-514. doi: 10.1111/plb.12554. Epub 2017 Mar 5. Plant Biol (Stuttg). 2017. PMID: 28145619
-
The Evolution of Sexual Fluids in Gymnosperms From Pollination Drops to Nectar.Front Plant Sci. 2018 Dec 18;9:1844. doi: 10.3389/fpls.2018.01844. eCollection 2018. Front Plant Sci. 2018. PMID: 30619413 Free PMC article. Review.
Cited by
-
Convergent evolution of fern nectaries facilitated independent recruitment of ant-bodyguards from flowering plants.Nat Commun. 2024 May 24;15(1):4392. doi: 10.1038/s41467-024-48646-x. Nat Commun. 2024. PMID: 38789437 Free PMC article.
References
-
- Abraham E., del Valle H. F., Roig F., Torres L., Ares J. O., Coronato F., et al. (2009). Overview of the geography of the Monte Desert biome (Argentina). J. Arid Environ. 73 144–153. 10.1016/j.jaridenv.2008.09.028 - DOI
-
- Aranda-Rickert A., Fracchia S. (2011a). Are subordinate ants the best seed dispersers? Linking dominance hierarchies and seed dispersal ability in myrmecochory interactions. Arthropod Plant Interact. 6 297–306. 10.1007/s11829-011-9166-z - DOI
-
- Aranda-Rickert A., Fracchia S. (2011b). Pogonomyrmex cunicularius as the keystone disperser of elaiosome-bearing Jatropha excisa seeds in semi-arid Argentina. Entomol. Exp. Appl. 139 91–102. 10.1111/j.1570-7458.2011.01111.x - DOI
-
- Bartoń K. (2019). MuMIn: multi-model inference. Available online at: https://CRAN.R-project.org/package=MuMIn (accessed April 15, 2020).
LinkOut - more resources
Full Text Sources
Miscellaneous