

p27Kip1, an Intrinsically Unstructured Protein with Scaffold Properties
- PMID: 34571903
- PMCID: PMC8465030
- DOI: 10.3390/cells10092254
p27Kip1, an Intrinsically Unstructured Protein with Scaffold Properties
Abstract
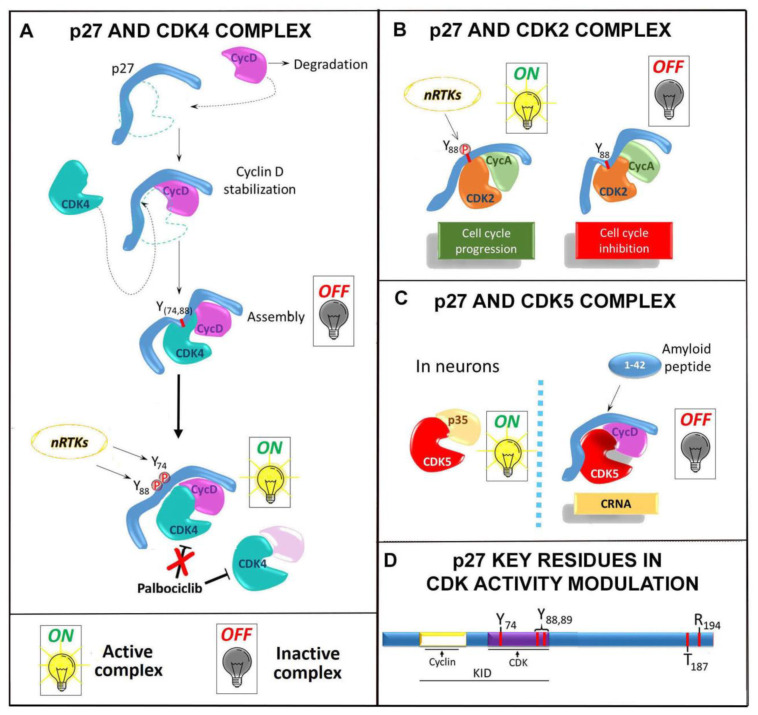
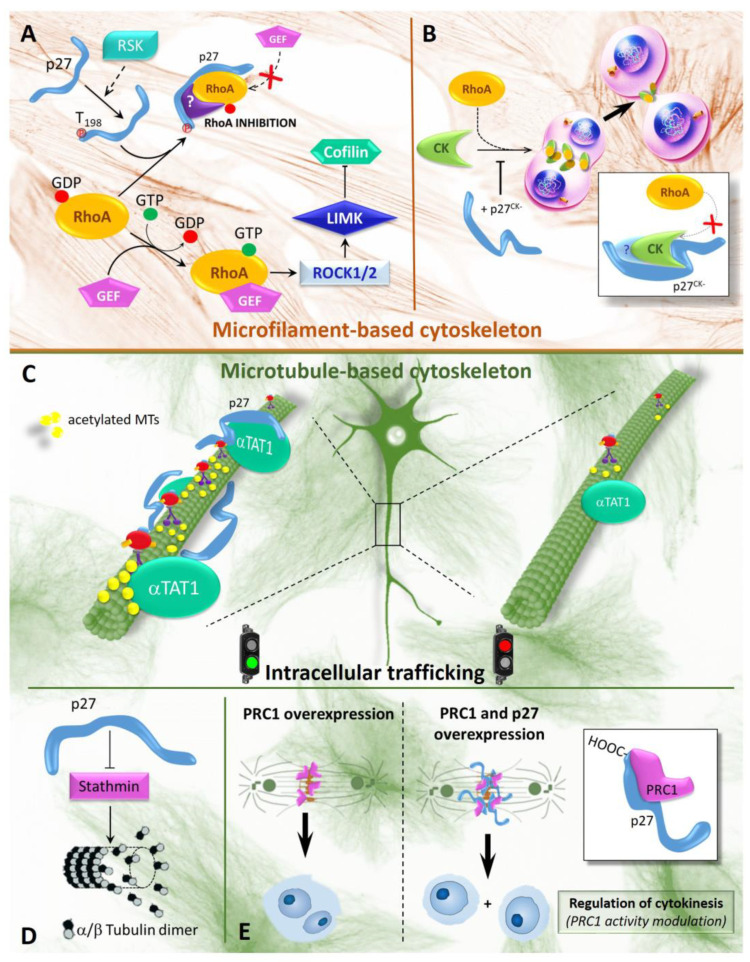
The Cyclin-dependent kinase (CDK) regulator p27Kip1 is a gatekeeper of G1/S transition. It also regulates G2/M progression and cytokinesis completion, via CDK-dependent or -independent mechanisms. Recently, other important p27Kip1 functions have been described, including the regulation of cell motility and migration, the control of cell differentiation program and the activation of apoptosis/autophagy. Several factors modulate p27Kip1 activities, including its level, cellular localization and post-translational modifications. As a matter of fact, the protein is phosphorylated, ubiquitinated, SUMOylated, O-linked N-acetylglicosylated and acetylated on different residues. p27Kip1 belongs to the family of the intrinsically unstructured proteins and thus it is endowed with a large flexibility and numerous interactors, only partially identified. In this review, we look at p27Kip1 properties and ascribe part of its heterogeneous functions to the ability to act as an anchor or scaffold capable to participate in the construction of different platforms for modulating cell response to extracellular signals and allowing adaptation to environmental changes.
Keywords: CDK; Rho GTPase; cyclin; cytoskeleton; intrinsically unstructured protein; p27Kip1; scaffold protein; αTAT1.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Newton I.S. In: Philosophiae Naturalis Principia Mathematica. 1st ed. Streater J., editor. Royal Society of London; London, UK: 1687.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
