

Impact of G-Quadruplexes on the Regulation of Genome Integrity, DNA Damage and Repair
- PMID: 34572497
- PMCID: PMC8472537
- DOI: 10.3390/biom11091284
Impact of G-Quadruplexes on the Regulation of Genome Integrity, DNA Damage and Repair
Abstract
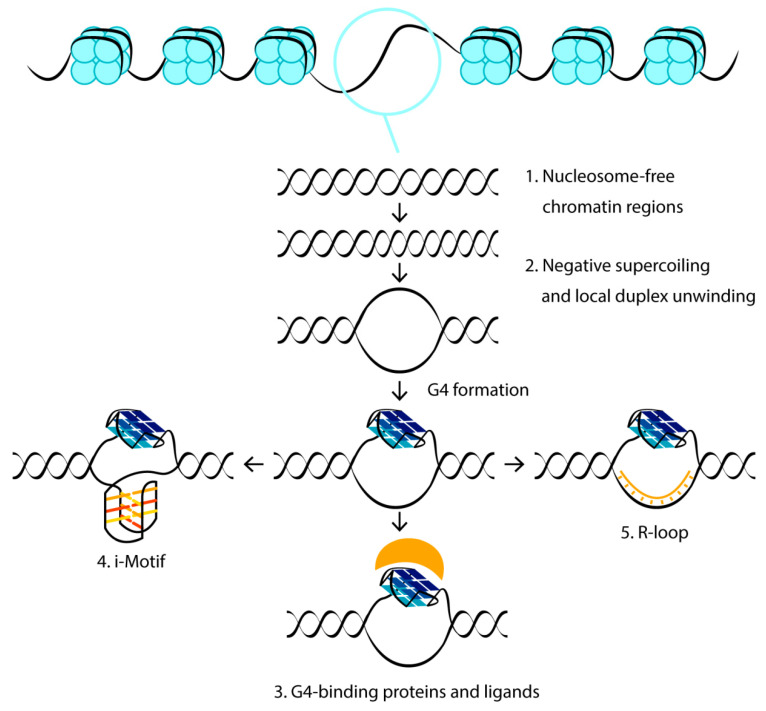
DNA G-quadruplexes (G4s) are known to be an integral part of the complex regulatory systems in both normal and pathological cells. At the same time, the ability of G4s to impede DNA replication plays a critical role in genome integrity. This review summarizes the results of recent studies of G4-mediated genomic and epigenomic instability, together with associated DNA damage and repair processes. Although the underlying mechanisms remain to be elucidated, it is known that, among the proteins that recognize G4 structures, many are linked to DNA repair. We analyzed the possible role of G4s in promoting double-strand DNA breaks, one of the most deleterious DNA lesions, and their repair via error-prone mechanisms. The patterns of G4 damage, with a focus on the introduction of oxidative guanine lesions, as well as their removal from G4 structures by canonical repair pathways, were also discussed together with the effects of G4s on the repair machinery. According to recent findings, there must be a delicate balance between G4-induced genome instability and G4-promoted repair processes. A broad overview of the factors that modulate the stability of G4 structures in vitro and in vivo is also provided here.
Keywords: DNA damage; DNA repair; G-quadruplex; G-quadruplex-binding proteins; genomic instability.
Conflict of interest statement
The authors declare no conflict of interest.
Figures






References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
