

TAR RNA Mediated Folding of a Single-Arginine-Mutant HIV-1 Tat Protein within HeLa Cells Experiencing Intracellular Crowding
- PMID: 34576162
- PMCID: PMC8468913
- DOI: 10.3390/ijms22189998
TAR RNA Mediated Folding of a Single-Arginine-Mutant HIV-1 Tat Protein within HeLa Cells Experiencing Intracellular Crowding
Abstract
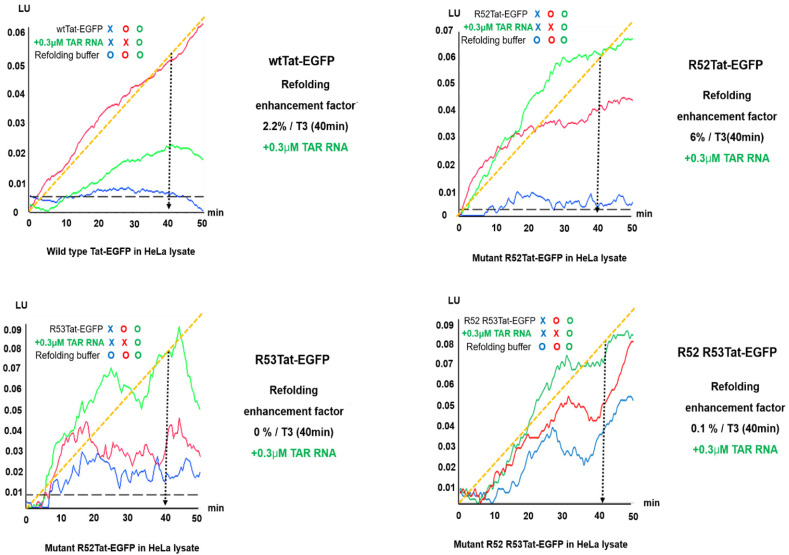
The various effects of native protein folding on the stability and folding rate of intrinsically disordered proteins (IDPs) in crowded intracellular environments are important in biomedicine. Although most studies on protein folding have been conducted in vitro, providing valuable insights, studies on protein folding in crowded intracellular environments are scarce. This study aimed to explore the effects of intracellular molecular crowding on the folding of mutant transactivator HIV-1 Tat based on intracellular interactions, including TAR RNA, as proof of the previously reported chaperna-RNA concept. Considering that the Tat-TAR RNA motif binds RNA, we assessed the po tential function of TAR RNA as a chaperna for the refolding of R52Tat, a mutant in which the argi nine (R) residues at R52 have been replaced with alanine (A) by site-directed mutagenesis. We mon itored Tat-EGFP and Tat folding in HeLa cells via time-lapse fluorescence microscopy and biolayer interferometry using EGFP fusion as an indicator for folding status. These results show that the refolding of R52A Tat was stimulated well at a 0.3 μM TAR RNA concentration; wild-type Tat refolding was essentially abolished because of a reduction in the affinity for TAR RNA at that con centration. The folding and refolding of R52Tat were mainly promoted upon stimulation with TAR RNA. Our findings provide novel insights into the therapeutic potential of chaperna-mediated fold ing through the examination of as-yet-unexplored RNA-mediated protein folding as well as viral genetic variants that modulate viral evolutionary linkages for viral diseases inside a crowded intra cellular environment.
Keywords: TAR RNA; Tat; arginine-rich domains; chaperna; crowding effects; folding kinetics.
Conflict of interest statement
The authors declare no conflict of interest.
Figures




Similar articles
-
The folding competence of HIV-1 Tat mediated by interaction with TAR RNA.RNA Biol. 2017 Jul 3;14(7):926-937. doi: 10.1080/15476286.2017.1311455. Epub 2017 Apr 18. RNA Biol. 2017. PMID: 28418268 Free PMC article.
-
The number of positively charged amino acids in the basic domain of Tat is critical for trans-activation and complex formation with TAR RNA.Proc Natl Acad Sci U S A. 1991 Jul 15;88(14):6234-8. doi: 10.1073/pnas.88.14.6234. Proc Natl Acad Sci U S A. 1991. PMID: 2068104 Free PMC article.
-
High affinity binding of TAR RNA by the human immunodeficiency virus type-1 tat protein requires base-pairs in the RNA stem and amino acid residues flanking the basic region.J Mol Biol. 1993 Mar 5;230(1):90-110. doi: 10.1006/jmbi.1993.1128. J Mol Biol. 1993. PMID: 8450553
-
Face-time with TAR: Portraits of an HIV-1 RNA with diverse modes of effector recognition relevant for drug discovery.J Biol Chem. 2019 Jun 14;294(24):9326-9341. doi: 10.1074/jbc.REV119.006860. Epub 2019 May 12. J Biol Chem. 2019. PMID: 31080171 Free PMC article. Review.
-
Discoveries of Tat-TAR interaction inhibitors for HIV-1.Curr Drug Targets Infect Disord. 2005 Dec;5(4):433-44. doi: 10.2174/156800505774912901. Curr Drug Targets Infect Disord. 2005. PMID: 16535863 Review.
Cited by
-
A Novel Time-Resolved Fluorescence Resonance Energy Transfer Assay for the Discovery of Small-Molecule Inhibitors of HIV-1 Tat-Regulated Transcription.Int J Mol Sci. 2023 May 23;24(11):9139. doi: 10.3390/ijms24119139. Int J Mol Sci. 2023. PMID: 37298089 Free PMC article.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
