

A prenylated dsRNA sensor protects against severe COVID-19
- PMID: 34581622
- PMCID: PMC7612834
- DOI: 10.1126/science.abj3624
A prenylated dsRNA sensor protects against severe COVID-19
Abstract
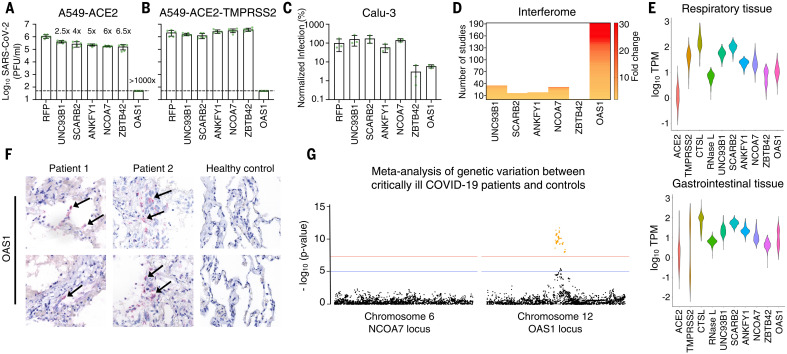
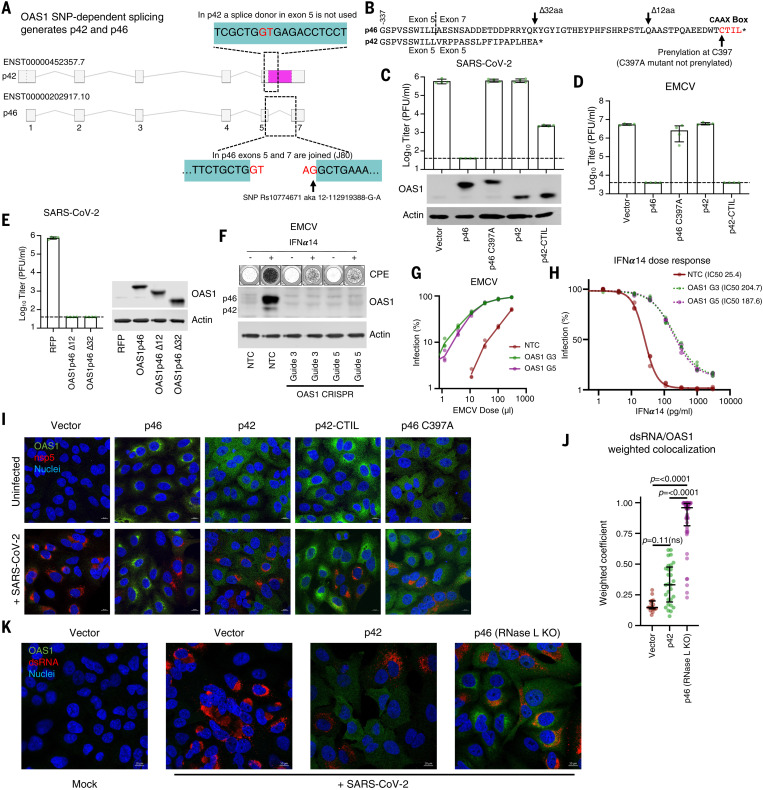
Inherited genetic factors can influence the severity of COVID-19, but the molecular explanation underpinning a genetic association is often unclear. Intracellular antiviral defenses can inhibit the replication of viruses and reduce disease severity. To better understand the antiviral defenses relevant to COVID-19, we used interferon-stimulated gene (ISG) expression screening to reveal that 2′-5′-oligoadenylate synthetase 1 (OAS1), through ribonuclease L, potently inhibits severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2). We show that a common splice-acceptor single-nucleotide polymorphism (Rs10774671) governs whether patients express prenylated OAS1 isoforms that are membrane-associated and sense-specific regions of SARS-CoV-2 RNAs or if they only express cytosolic, nonprenylated OAS1 that does not efficiently detect SARS-CoV-2. In hospitalized patients, expression of prenylated OAS1 was associated with protection from severe COVID-19, suggesting that this antiviral defense is a major component of a protective antiviral response.
Figures





Comment in
-
Defective viral RNA sensing linked to severe COVID-19.Science. 2021 Oct 29;374(6567):535-536. doi: 10.1126/science.abm3921. Epub 2021 Oct 28. Science. 2021. PMID: 34709914
References
-
- Pairo-Castineira E., Clohisey S., Klaric L., Bretherick A. D., Rawlik K., Pasko D., Walker S., Parkinson N., Fourman M. H., Russell C. D., Furniss J., Richmond A., Gountouna E., Wrobel N., Harrison D., Wang B., Wu Y., Meynert A., Griffiths F., Oosthuyzen W., Kousathanas A., Moutsianas L., Yang Z., Zhai R., Zheng C., Grimes G., Beale R., Millar J., Shih B., Keating S., Zechner M., Haley C., Porteous D. J., Hayward C., Yang J., Knight J., Summers C., Shankar-Hari M., Klenerman P., Turtle L., Ho A., Moore S. C., Hinds C., Horby P., Nichol A., Maslove D., Ling L., McAuley D., Montgomery H., Walsh T., Pereira A. C., Renieri A., Shen X., Ponting C. P., Fawkes A., Tenesa A., Caulfield M., Scott R., Rowan K., Murphy L., Openshaw P. J. M., Semple M. G., Law A., Vitart V., Wilson J. F., Baillie J. K.; GenOMICC Investigators; ISARIC4C Investigators; COVID-19 Human Genetics Initiative; 23andMe Investigators; BRACOVID Investigators; Gen-COVID Investigators , Genetic mechanisms of critical illness in COVID-19. Nature 591, 92–98 (2021). 10.1038/s41586-020-03065-y - DOI - PubMed
-
- Bastard P., Rosen L. B., Zhang Q., Michailidis E., Hoffmann H.-H., Zhang Y., Dorgham K., Philippot Q., Rosain J., Béziat V., Manry J., Shaw E., Haljasmägi L., Peterson P., Lorenzo L., Bizien L., Trouillet-Assant S., Dobbs K., de Jesus A. A., Belot A., Kallaste A., Catherinot E., Tandjaoui-Lambiotte Y., Le Pen J., Kerner G., Bigio B., Seeleuthner Y., Yang R., Bolze A., Spaan A. N., Delmonte O. M., Abers M. S., Aiuti A., Casari G., Lampasona V., Piemonti L., Ciceri F., Bilguvar K., Lifton R. P., Vasse M., Smadja D. M., Migaud M., Hadjadj J., Terrier B., Duffy D., Quintana-Murci L., van de Beek D., Roussel L., Vinh D. C., Tangye S. G., Haerynck F., Dalmau D., Martinez-Picado J., Brodin P., Nussenzweig M. C., Boisson-Dupuis S., Rodríguez-Gallego C., Vogt G., Mogensen T. H., Oler A. J., Gu J., Burbelo P. D., Cohen J. I., Biondi A., Bettini L. R., D’Angio M., Bonfanti P., Rossignol P., Mayaux J., Rieux-Laucat F., Husebye E. S., Fusco F., Ursini M. V., Imberti L., Sottini A., Paghera S., Quiros-Roldan E., Rossi C., Castagnoli R., Montagna D., Licari A., Marseglia G. L., Duval X., Ghosn J., Tsang J. S., Goldbach-Mansky R., Kisand K., Lionakis M. S., Puel A., Zhang S.-Y., Holland S. M., Gorochov G., Jouanguy E., Rice C. M., Cobat A., Notarangelo L. D., Abel L., Su H. C., Casanova J.-L.; HGID Lab; NIAID-USUHS Immune Response to COVID Group; COVID Clinicians; COVID-STORM Clinicians; Imagine COVID Group; French COVID Cohort Study Group; Milieu Intérieur Consortium; CoV-Contact Cohort; Amsterdam UMC Covid-19 Biobank; COVID Human Genetic Effort , Autoantibodies against type I IFNs in patients with life-threatening COVID-19. Science 370, eabd4585 (2020). 10.1126/science.abd4585 - DOI - PMC - PubMed
-
- Monk P. D., Marsden R. J., Tear V. J., Brookes J., Batten T. N., Mankowski M., Gabbay F. J., Davies D. E., Holgate S. T., Ho L.-P., Clark T., Djukanovic R., Wilkinson T. M. A.; Inhaled Interferon Beta COVID-19 Study Group , Safety and efficacy of inhaled nebulised interferon beta-1a (SNG001) for treatment of SARS-CoV-2 infection: A randomised, double-blind, placebo-controlled, phase 2 trial. Lancet Respir. Med. 9, 196–206 (2021). 10.1016/S2213-2600(20)30511-7 - DOI - PMC - PubMed
-
- Channappanavar R., Fehr A. R., Vijay R., Mack M., Zhao J., Meyerholz D. K., Perlman S., Dysregulated type I interferon and inflammatory monocyte-macrophage responses cause lethal pneumonia in SARS-CoV-infected mice. Cell Host Microbe 19, 181–193 (2016). 10.1016/j.chom.2016.01.007 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- MC_PC_20060/MRC_/Medical Research Council/United Kingdom
- MR/V028448/1/MRC_/Medical Research Council/United Kingdom
- MR/R021562/1/MRC_/Medical Research Council/United Kingdom
- MC_PC_19025/MRC_/Medical Research Council/United Kingdom
- MC_UU_12014/12/MRC_/Medical Research Council/United Kingdom
- G0701652/MRC_/Medical Research Council/United Kingdom
- MR/S00971X/1/MRC_/Medical Research Council/United Kingdom
- MC_UU_12014/10/MRC_/Medical Research Council/United Kingdom
- MR/K024752/1/MRC_/Medical Research Council/United Kingdom
- MR/P022642/1/MRC_/Medical Research Council/United Kingdom
- MR/V000489/1/MRC_/Medical Research Council/United Kingdom
- CO-CIN-01/DH_/Department of Health/United Kingdom
- MC_UU_12014/9/MRC_/Medical Research Council/United Kingdom
- 204870/Z/16/Z/WT_/Wellcome Trust/United Kingdom
- MC_UU_12014/3/MRC_/Medical Research Council/United Kingdom
- IS-BRC-1215-20013/DH_/Department of Health/United Kingdom
- MR/V011561/1/MRC_/Medical Research Council/United Kingdom
- 201366/WT_/Wellcome Trust/United Kingdom
- 210688/Z/18/Z/WT_/Wellcome Trust/United Kingdom
- 209412/WT_/Wellcome Trust/United Kingdom
- MR/S020039/1/MRC_/Medical Research Council/United Kingdom
- BB/M011224/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- 201366/Z/16/Z/WT_/Wellcome Trust/United Kingdom
- MC_PC_19026/MRC_/Medical Research Council/United Kingdom
- 210688/WT_/Wellcome Trust/United Kingdom
- MR/P001602/1/MRC_/Medical Research Council/United Kingdom
- MC_PC_19059/MRC_/Medical Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
