

The human sperm basal body is a complex centrosome important for embryo preimplantation development
- PMID: 34581808
- PMCID: PMC8561016
- DOI: 10.1093/molehr/gaab062
The human sperm basal body is a complex centrosome important for embryo preimplantation development
Abstract
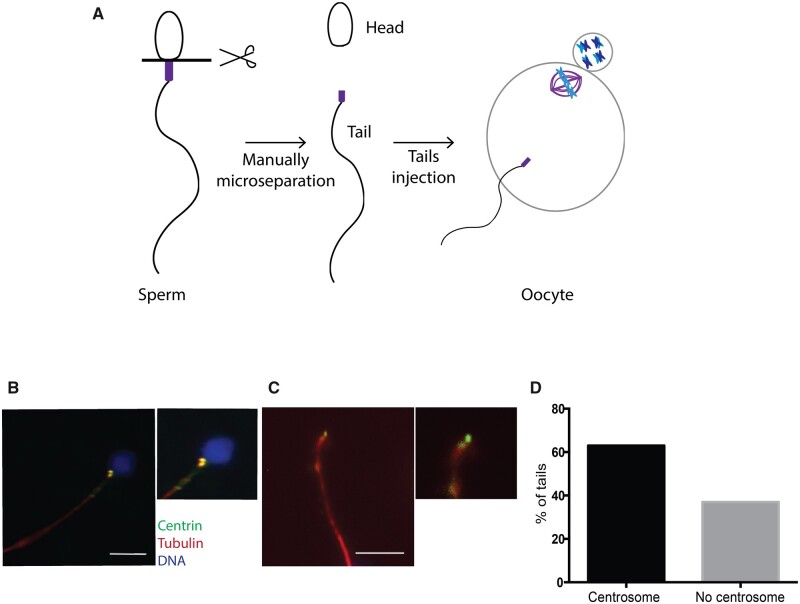
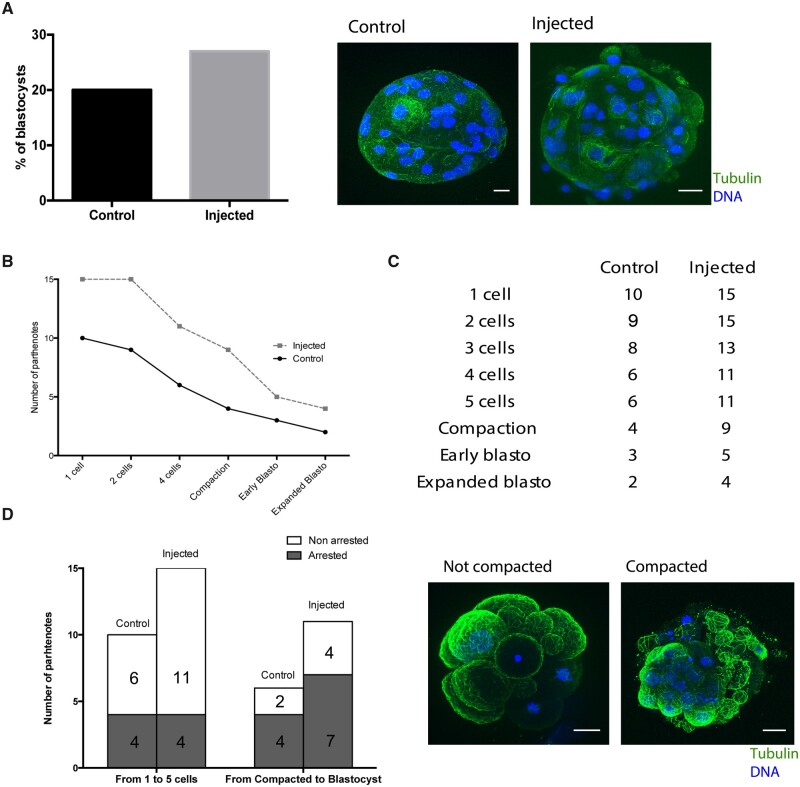
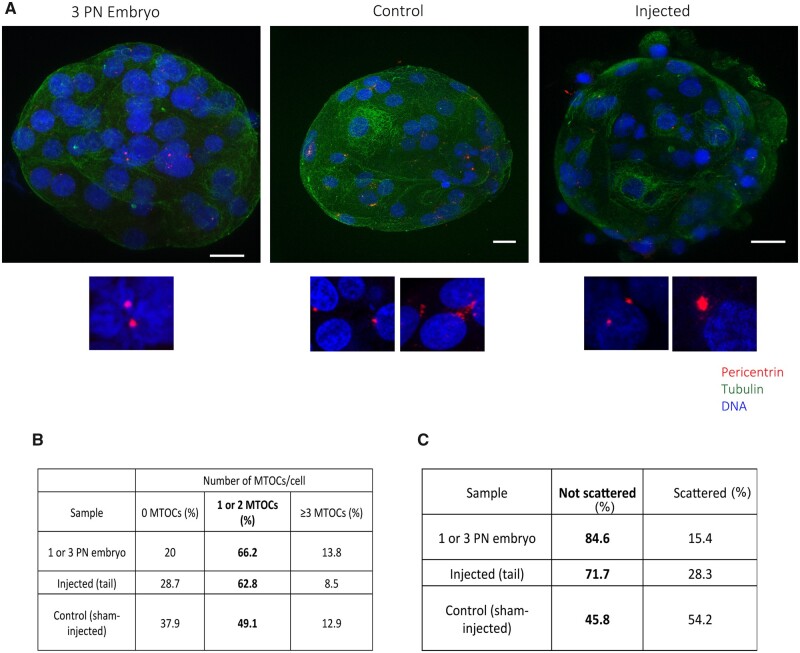
The mechanism of conversion of the human sperm basal body to a centrosome after fertilization, and its role in supporting human early embryogenesis, has not been directly addressed so far. Using proteomics and immunofluorescence studies, we show here that the human zygote inherits a basal body enriched with centrosomal proteins from the sperm, establishing the first functional centrosome of the new organism. Injection of human sperm tails containing the basal body into human oocytes followed by parthenogenetic activation, showed that the centrosome contributes to the robustness of the early cell divisions, increasing the probability of parthenotes reaching the compaction stage. In the absence of the sperm-derived centrosome, pericentriolar material (PCM) components stored in the oocyte can form de novo structures after genome activation, suggesting a tight PCM expression control in zygotes. Our results reveal that the sperm basal body is a complex organelle which converts to a centrosome after fertilization, ensuring the early steps of embryogenesis and successful compaction. However, more experiments are needed to elucidate the exact molecular mechanisms of centrosome inheritance in humans.
Keywords: centriole; centrosome; compaction; embryo early development; fertilization; human; microtubule organizing centers; pericentriolar material.
© The Author(s) 2021. Published by Oxford University Press on behalf of European Society of Human Reproduction and Embryology.
Figures
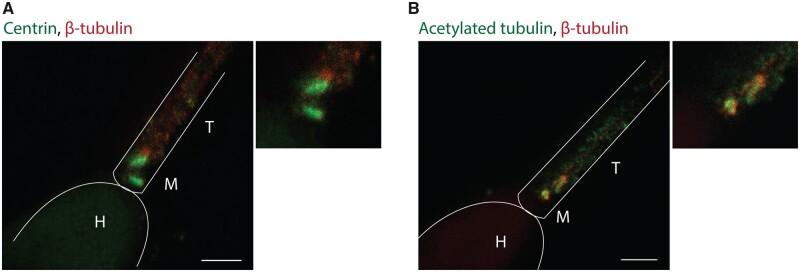
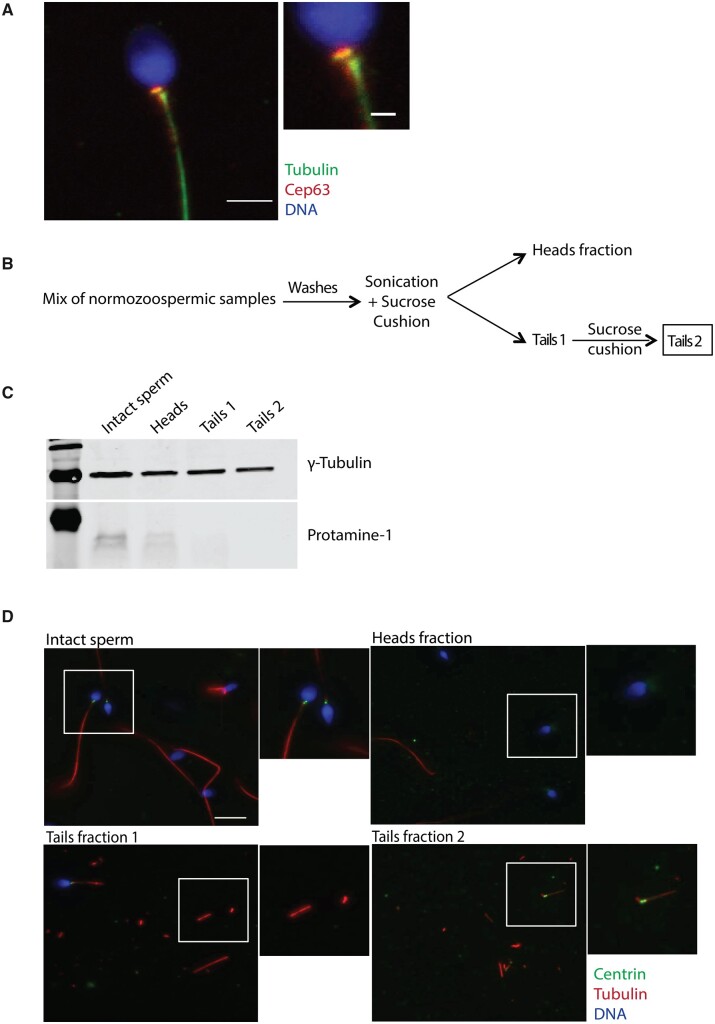
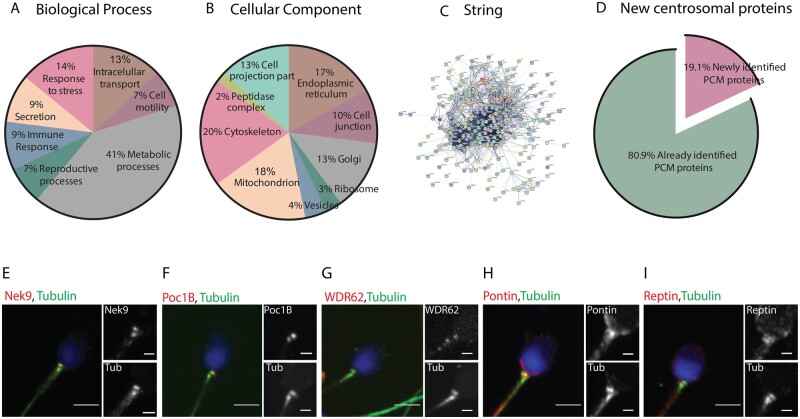
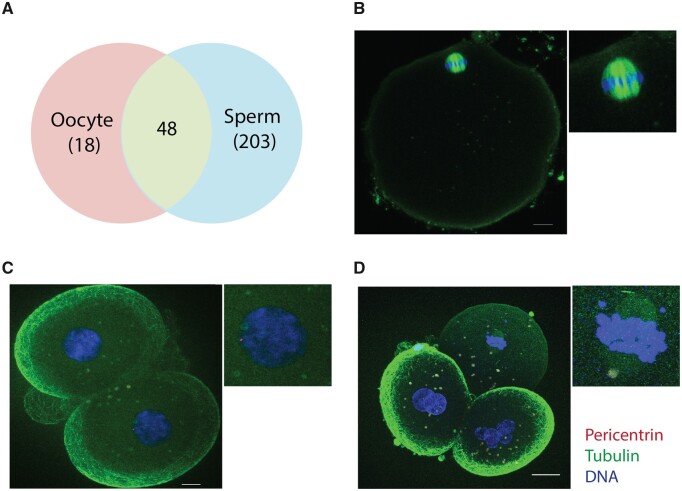
Similar articles
-
Characterization of human gamete centrosomes for assisted reproduction.Ital J Anat Embryol. 2001;106(2 Suppl 2):61-73. Ital J Anat Embryol. 2001. PMID: 11732597 Review.
-
Mitosis in the human embryo: the vital role of the sperm centrosome (centriole).Histol Histopathol. 1997 Jul;12(3):827-56. Histol Histopathol. 1997. PMID: 9225167 Review.
-
Loss and Rebirth of the Animal Microtubule Organizing Center: How Maternal Expression of Centrosomal Proteins Cooperates with the Sperm Centriole in Zygotic Centrosome Reformation.Bioessays. 2018 Apr;40(4):e1700135. doi: 10.1002/bies.201700135. Epub 2018 Mar 9. Bioessays. 2018. PMID: 29522658
-
The sperm centriole: its inheritance, replication and perpetuation in early human embryos.Hum Reprod. 1996 Feb;11(2):345-56. doi: 10.1093/humrep/11.2.345. Hum Reprod. 1996. PMID: 8671223
-
Paternal contributions to the mammalian zygote: fertilization after sperm-egg fusion.Int Rev Cytol. 2000;195:1-65. doi: 10.1016/s0074-7696(08)62703-5. Int Rev Cytol. 2000. PMID: 10603574 Review.
Cited by
-
Profiling the male germline genome to unravel its reproductive potential.Fertil Steril. 2023 Feb;119(2):196-206. doi: 10.1016/j.fertnstert.2022.11.006. Epub 2022 Nov 12. Fertil Steril. 2023. PMID: 36379263 Free PMC article.
-
The Centriole's Role in Miscarriages.Front Cell Dev Biol. 2022 Mar 1;10:864692. doi: 10.3389/fcell.2022.864692. eCollection 2022. Front Cell Dev Biol. 2022. PMID: 35300410 Free PMC article.
-
The evolution of centriole degradation in mouse sperm.Nat Commun. 2024 Jan 2;15(1):117. doi: 10.1038/s41467-023-44411-8. Nat Commun. 2024. PMID: 38168044 Free PMC article.
-
Atypical Centriolar Composition Correlates with Internal Fertilization in Fish.Cells. 2022 Feb 22;11(5):758. doi: 10.3390/cells11050758. Cells. 2022. PMID: 35269380 Free PMC article. Review.
-
Chromosome Division in Early Embryos-Is Everything under Control? And Is the Cell Size Important?Int J Mol Sci. 2024 Feb 9;25(4):2101. doi: 10.3390/ijms25042101. Int J Mol Sci. 2024. PMID: 38396778 Free PMC article. Review.
References
-
- Amargant F, Barragan M, Vassena R, Vernos I.. Insights of the tubulin code in gametes and embryos: from basic research to potential clinical applications in humans. Biol Reprod 2019;100:575–589. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
