

In vivo CRISPR screens identify the E3 ligase Cop1 as a modulator of macrophage infiltration and cancer immunotherapy target
- PMID: 34582788
- PMCID: PMC9136996
- DOI: 10.1016/j.cell.2021.09.006
In vivo CRISPR screens identify the E3 ligase Cop1 as a modulator of macrophage infiltration and cancer immunotherapy target
Abstract
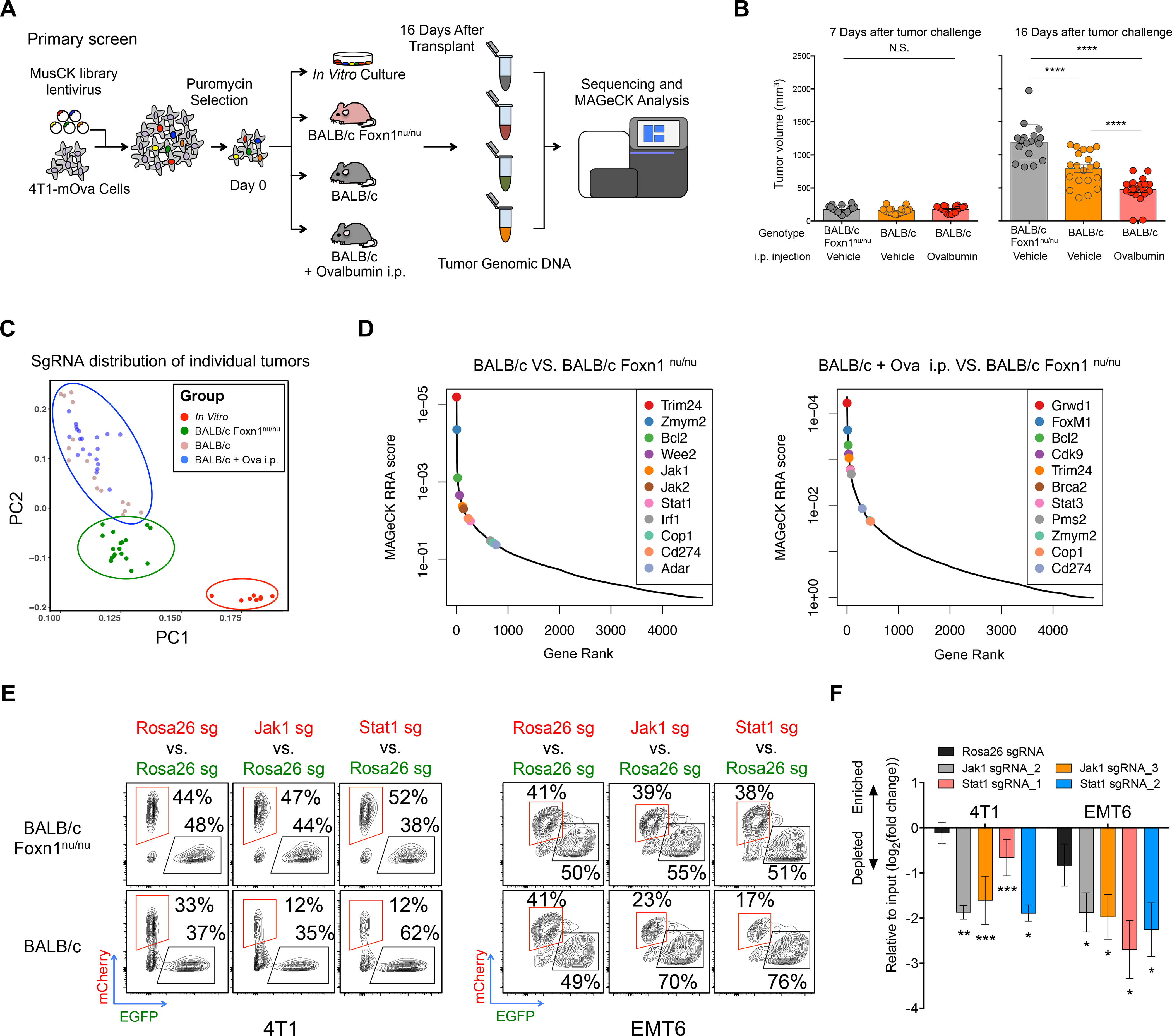
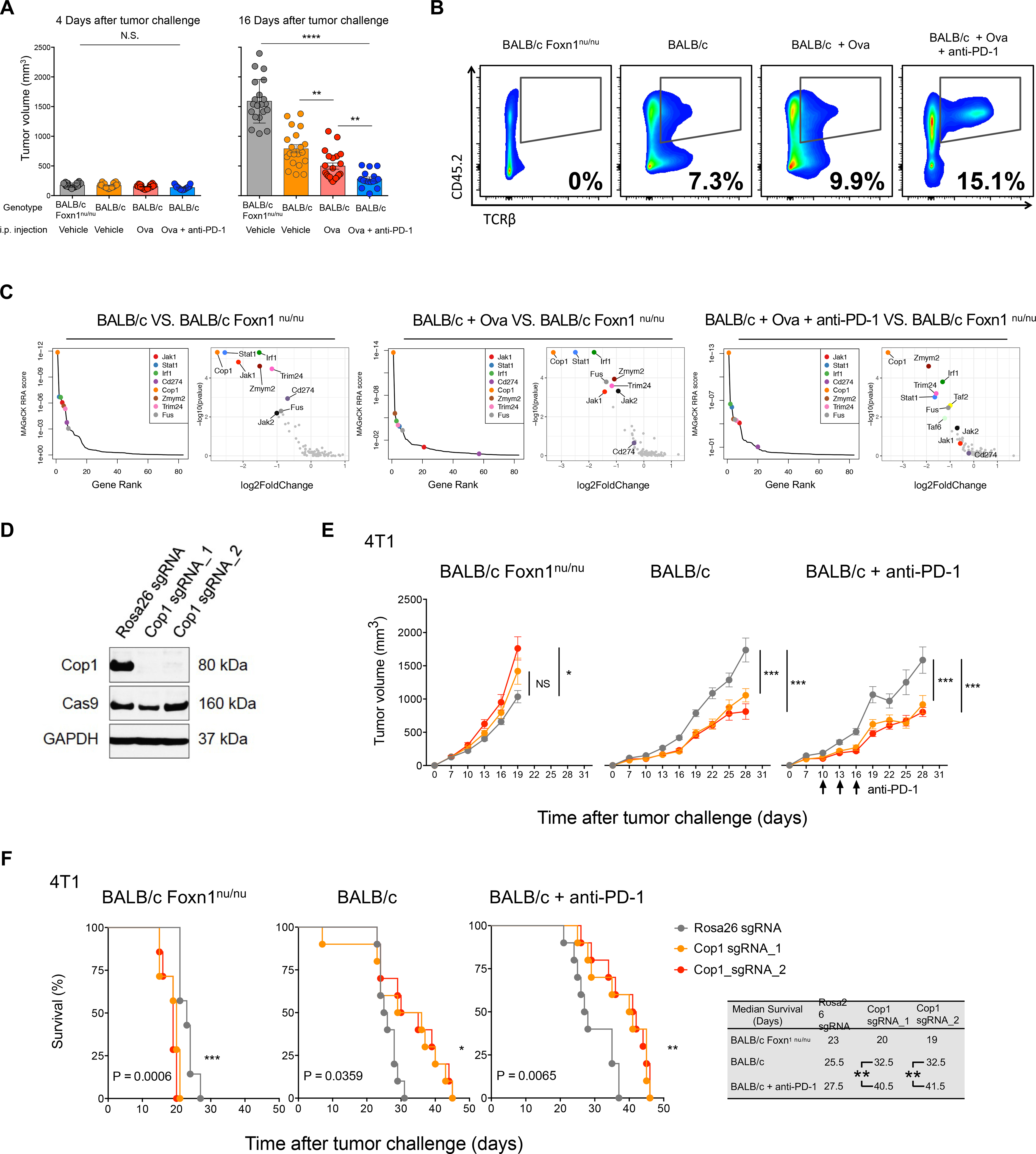
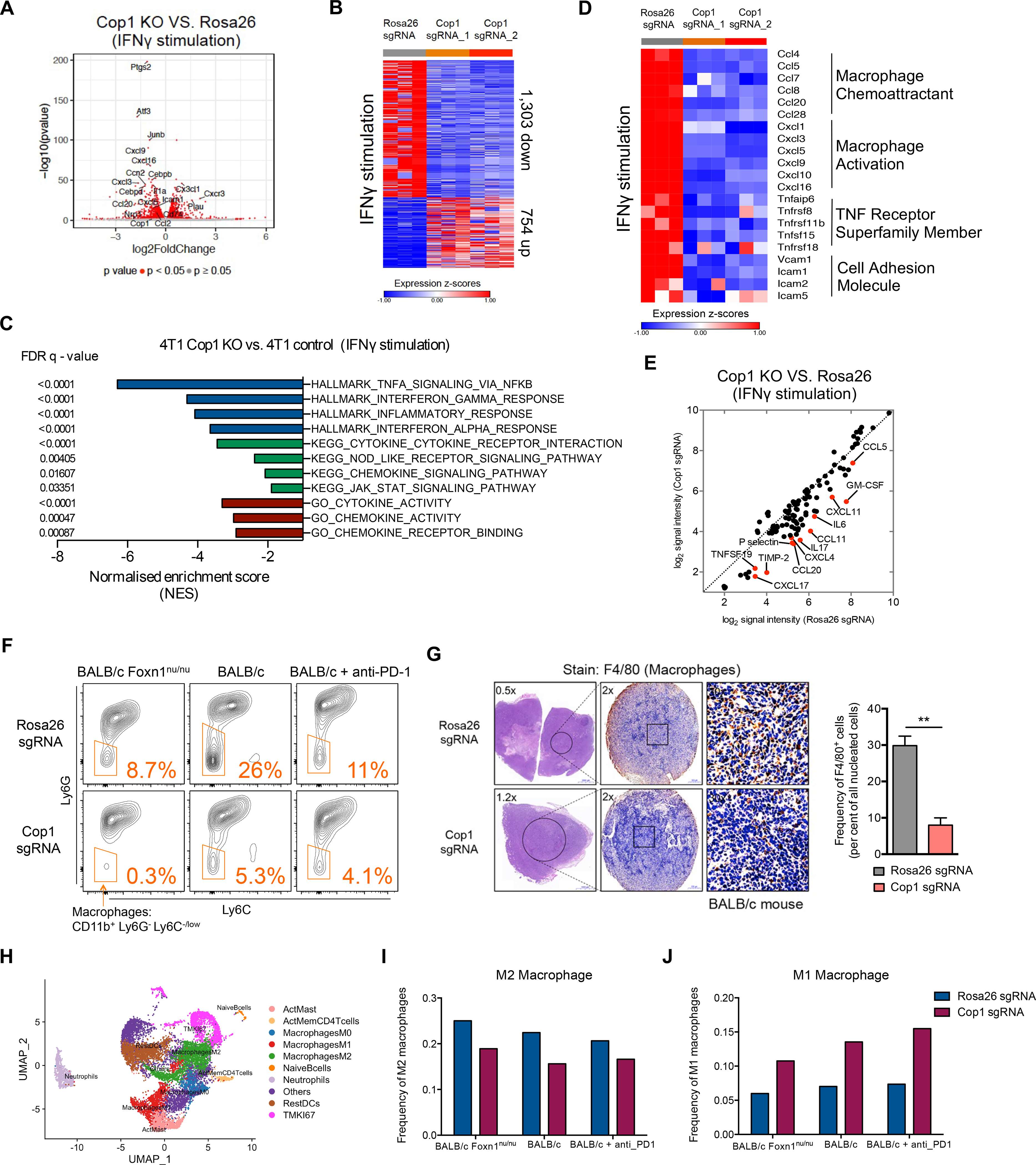
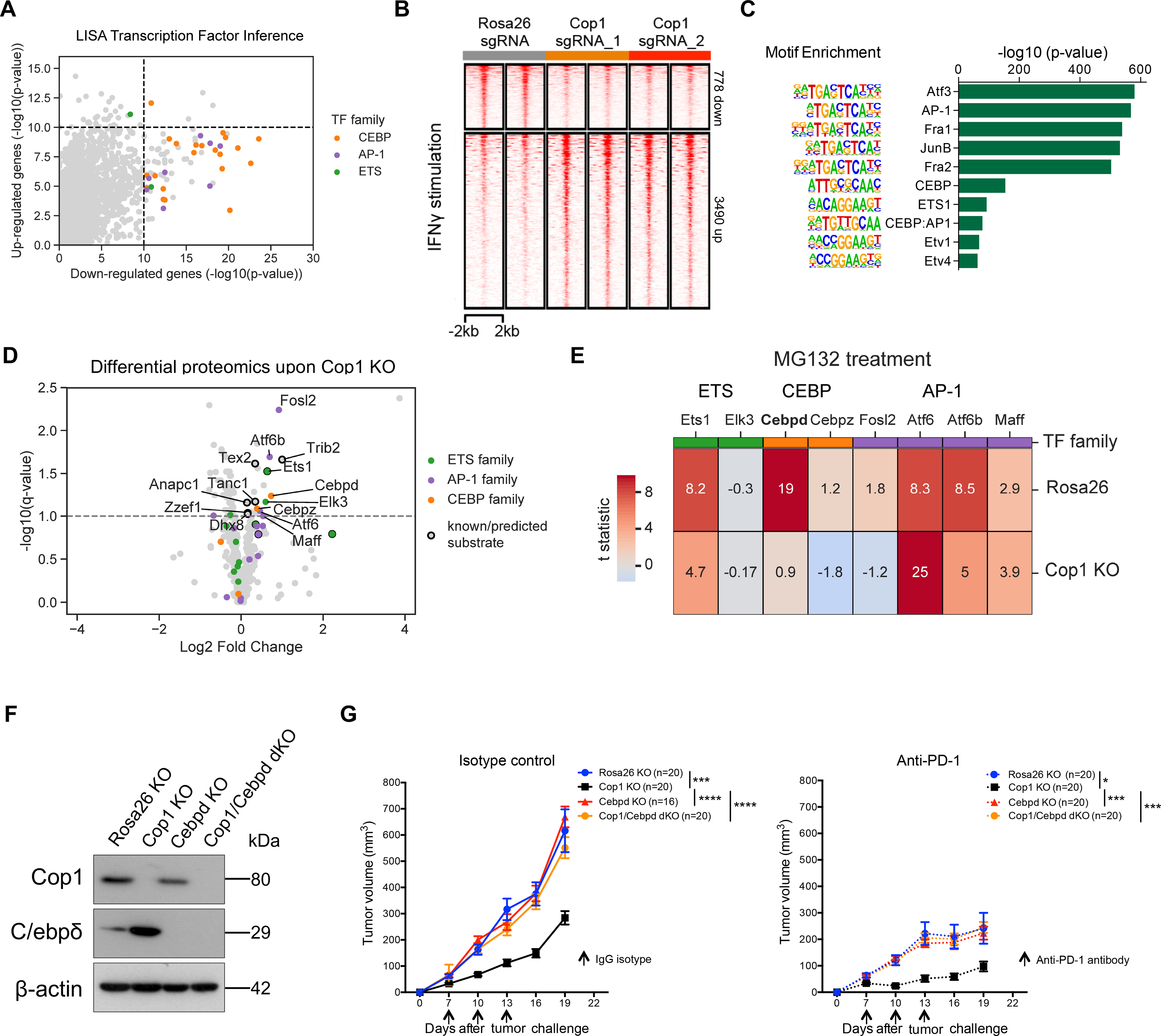
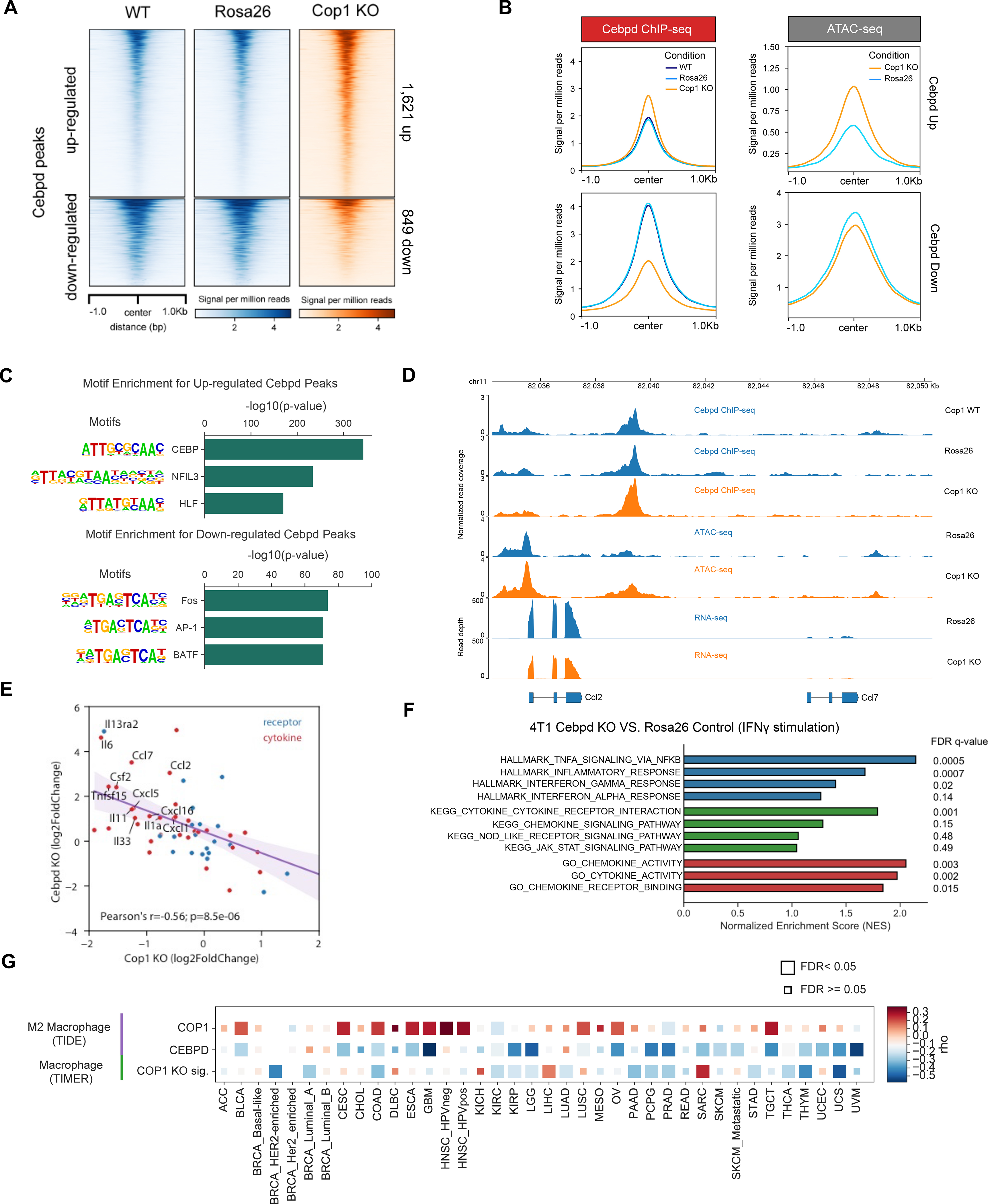
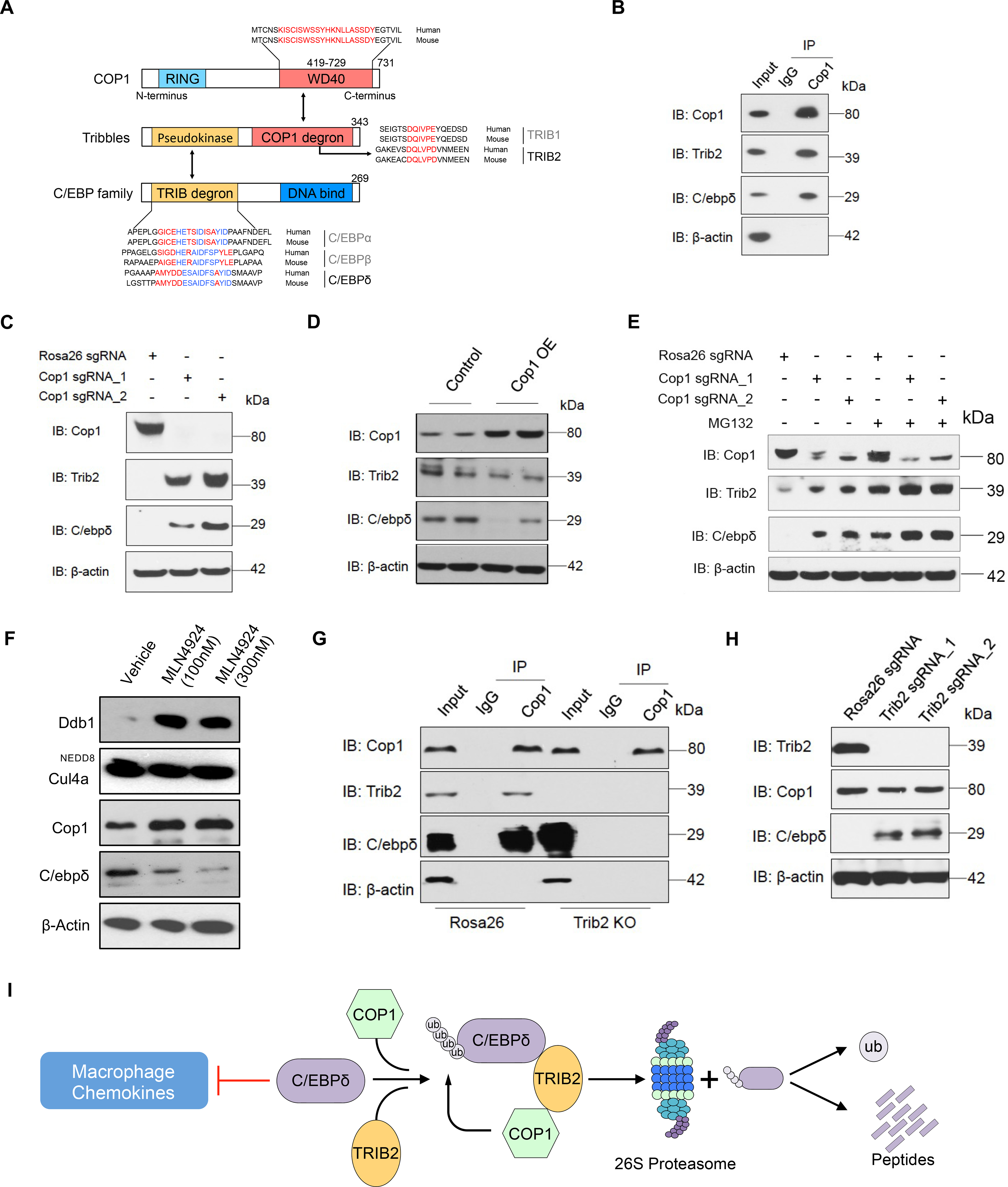
Despite remarkable clinical efficacy of immune checkpoint blockade (ICB) in cancer treatment, ICB benefits for triple-negative breast cancer (TNBC) remain limited. Through pooled in vivo CRISPR knockout (KO) screens in syngeneic TNBC mouse models, we found that deletion of the E3 ubiquitin ligase Cop1 in cancer cells decreases secretion of macrophage-associated chemokines, reduces tumor macrophage infiltration, enhances anti-tumor immunity, and strengthens ICB response. Transcriptomics, epigenomics, and proteomics analyses revealed that Cop1 functions through proteasomal degradation of the C/ebpδ protein. The Cop1 substrate Trib2 functions as a scaffold linking Cop1 and C/ebpδ, which leads to polyubiquitination of C/ebpδ. In addition, deletion of the E3 ubiquitin ligase Cop1 in cancer cells stabilizes C/ebpδ to suppress expression of macrophage chemoattractant genes. Our integrated approach implicates Cop1 as a target for improving cancer immunotherapy efficacy in TNBC by regulating chemokine secretion and macrophage infiltration in the tumor microenvironment.
Keywords: C/ebpδ; CRISPR screening; Cop1; E3 ubiquitin ligase; immunotherapy; triple-negative breast cancer.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests X.S.L. is a cofounder, board member, scientific advisor board member, and consultant of GV20 Oncotherapy and a stockholder of BMY, TMO, WBA, ABT, ABBV, and JNJ and receives research funding from Takeda, Sanofi, Bristol Myers Squibb, and Novartis. M.B. receives sponsored research support from and is a consultant to Novartis. M.B. serves on the scientific advisory boards of GV20 Oncotherapy, Kronos Bio, and H3 Biomedicine. T.X. is a cofounder, board member, and full-time employee of GV20 Oncotherapy.
Figures
References
-
- Beatty GL, and Paterson Y (2000). IFN-gamma can promote tumor evasion of the immune system in vivo by down-regulating cellular levels of an endogenous tumor antigen. J. Immunol. 165, 5502–5508. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
