

SATB2 preserves colon stem cell identity and mediates ileum-colon conversion via enhancer remodeling
- PMID: 34582804
- PMCID: PMC8741647
- DOI: 10.1016/j.stem.2021.09.004
SATB2 preserves colon stem cell identity and mediates ileum-colon conversion via enhancer remodeling
Abstract
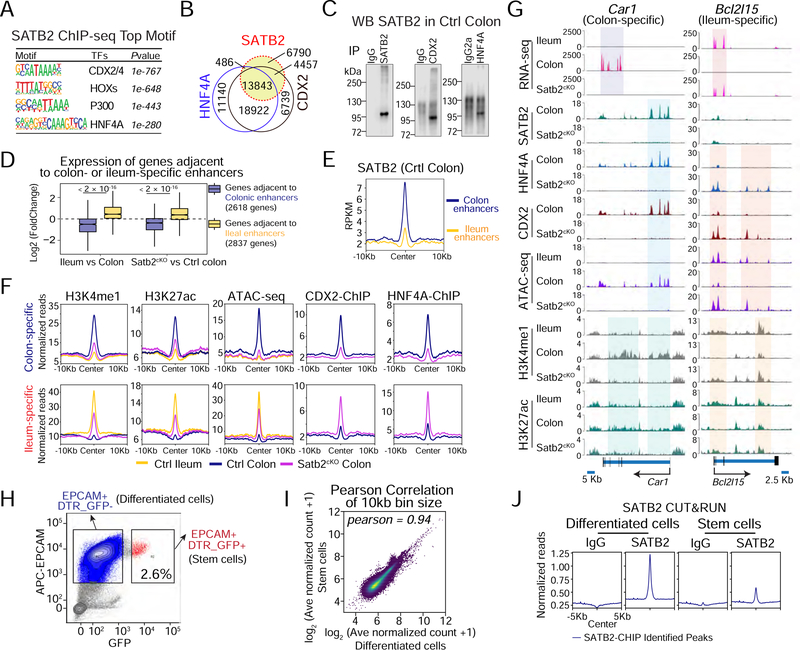
Adult stem cells maintain regenerative tissue structure and function by producing tissue-specific progeny, but the factors that preserve their tissue identities are not well understood. The small and large intestines differ markedly in cell composition and function, reflecting their distinct stem cell populations. Here we show that SATB2, a colon-restricted chromatin factor, singularly preserves LGR5+ adult colonic stem cell and epithelial identity in mice and humans. Satb2 loss in adult mice leads to stable conversion of colonic stem cells into small intestine ileal-like stem cells and replacement of the colonic mucosa with one that resembles the ileum. Conversely, SATB2 confers colonic properties on the mouse ileum. Human colonic organoids also adopt ileal characteristics upon SATB2 loss. SATB2 regulates colonic identity in part by modulating enhancer binding of the intestinal transcription factors CDX2 and HNF4A. Our study uncovers a conserved core regulator of colonic stem cells able to mediate cross-tissue plasticity in mature intestines.
Keywords: SATB2, intestine regeneration, colonic mucosa, stem cell conversion, enhancer remodelingintestine.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests A patent application related to this work is pending at Weill Cornell Medicine.
Figures






References
-
- Alcamo EA, Chirivella L, Dautzenberg M, Dobreva G, Farinas I, Grosschedl R, and McConnell SK (2008). Satb2 regulates callosal projection neuron identity in the developing cerebral cortex. Neuron 57, 364–377. - PubMed
-
- Barker N, van Es JH, Kuipers J, Kujala P, van den Born M, Cozijnsen M, Haegebarth A, Korving J, Begthel H, Peters PJ, et al. (2007). Identification of stem cells in small intestine and colon by marker gene Lgr5. Nature 449, 1003–1007. - PubMed
-
- Beumer J, and Clevers H (2021). Cell fate specification and differentiation in the adult mammalian intestine. Nat Rev Mol Cell Biol 22, 39–53. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
