

The missing enzymatic link in syntrophic methane formation from fatty acids
- PMID: 34583996
- PMCID: PMC8501807
- DOI: 10.1073/pnas.2111682118
The missing enzymatic link in syntrophic methane formation from fatty acids
Abstract
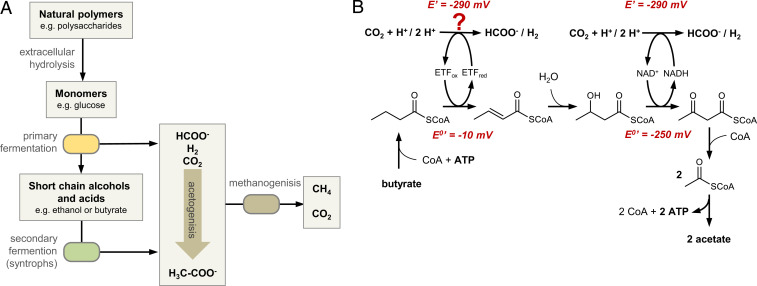
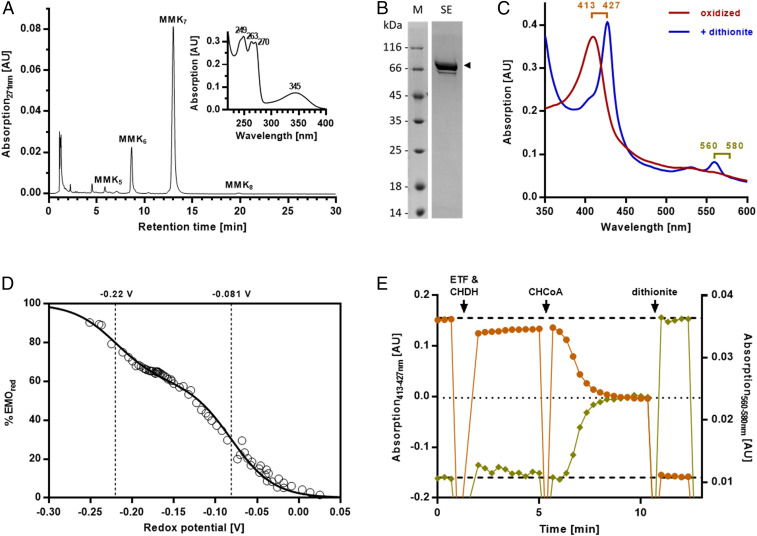
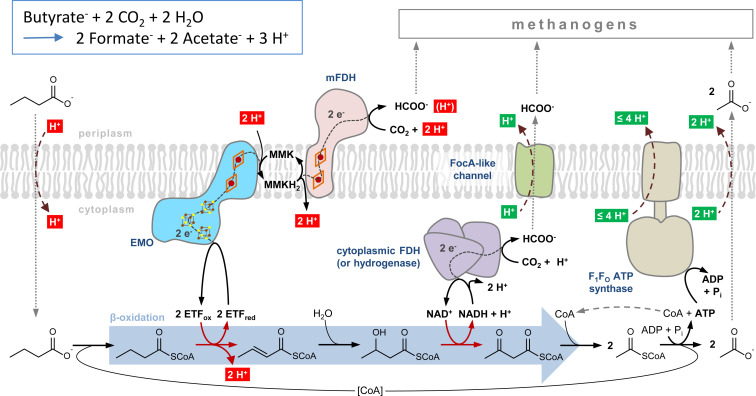
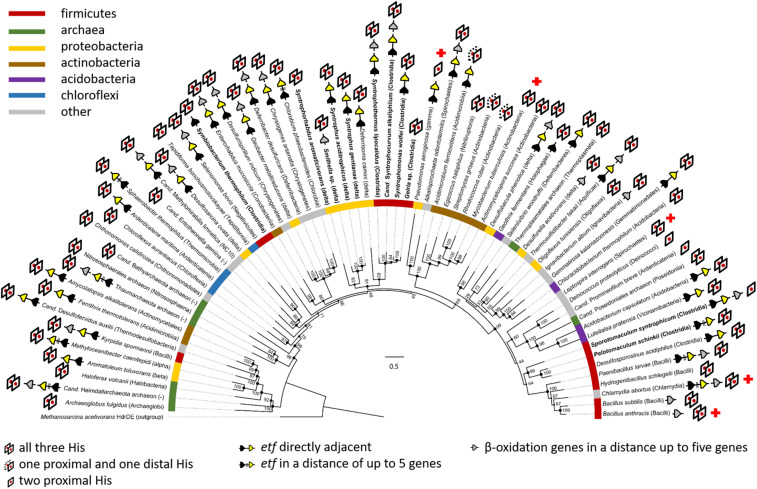
The microbial production of methane from organic matter is an essential process in the global carbon cycle and an important source of renewable energy. It involves the syntrophic interaction between methanogenic archaea and bacteria that convert primary fermentation products such as fatty acids to the methanogenic substrates acetate, H2, CO2, or formate. While the concept of syntrophic methane formation was developed half a century ago, the highly endergonic reduction of CO2 to methane by electrons derived from β-oxidation of saturated fatty acids has remained hypothetical. Here, we studied a previously noncharacterized membrane-bound oxidoreductase (EMO) from Syntrophus aciditrophicus containing two heme b cofactors and 8-methylmenaquinone as key redox components of the redox loop-driven reduction of CO2 by acyl-coenzyme A (CoA). Using solubilized EMO and proteoliposomes, we reconstituted the entire electron transfer chain from acyl-CoA to CO2 and identified the transfer from a high- to a low-potential heme b with perfectly adjusted midpoint potentials as key steps in syntrophic fatty acid oxidation. The results close our gap of knowledge in the conversion of biomass into methane and identify EMOs as key players of β-oxidation in (methyl)menaquinone-containing organisms.
Keywords: diheme oxidoreductase; methylmenaquinone; microbial methane formation; redox loop; syntrophy.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Evans P. N., et al. ., An evolving view of methane metabolism in the Archaea. Nat. Rev. Microbiol. 17, 219–232 (2019). - PubMed
-
- Thauer R. K., Kaster A. K., Seedorf H., Buckel W., Hedderich R., Methanogenic archaea: Ecologically relevant differences in energy conservation. Nat. Rev. Microbiol. 6, 579–591 (2008). - PubMed
-
- Stams A. J. M., Plugge C. M., Electron transfer in syntrophic communities of anaerobic bacteria and archaea. Nat. Rev. Microbiol. 7, 568–577 (2009). - PubMed
-
- Sieber J. R., Le H. M., McInerney M. J., The importance of hydrogen and formate transfer for syntrophic fatty, aromatic and alicyclic metabolism. Environ. Microbiol. 16, 177–188 (2014). - PubMed
-
- Schink B., Montag D., Keller A., Müller N., Hydrogen or formate: Alternative key players in methanogenic degradation. Environ. Microbiol. Rep. 9, 189–202 (2017). - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
Miscellaneous
