

Actin crosslinker competition and sorting drive emergent GUV size-dependent actin network architecture
- PMID: 34584211
- PMCID: PMC8478941
- DOI: 10.1038/s42003-021-02653-6
Actin crosslinker competition and sorting drive emergent GUV size-dependent actin network architecture
Abstract
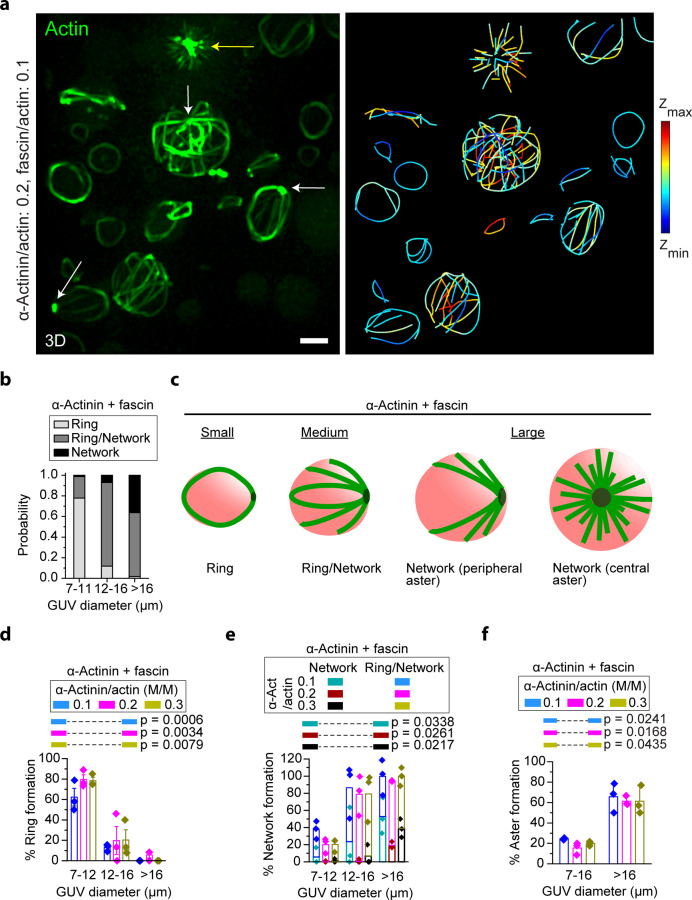
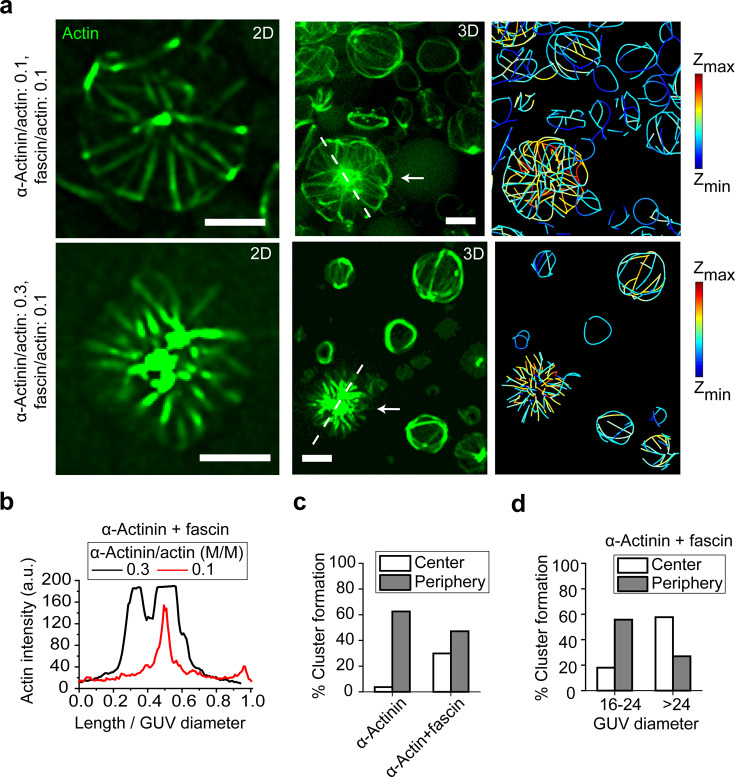
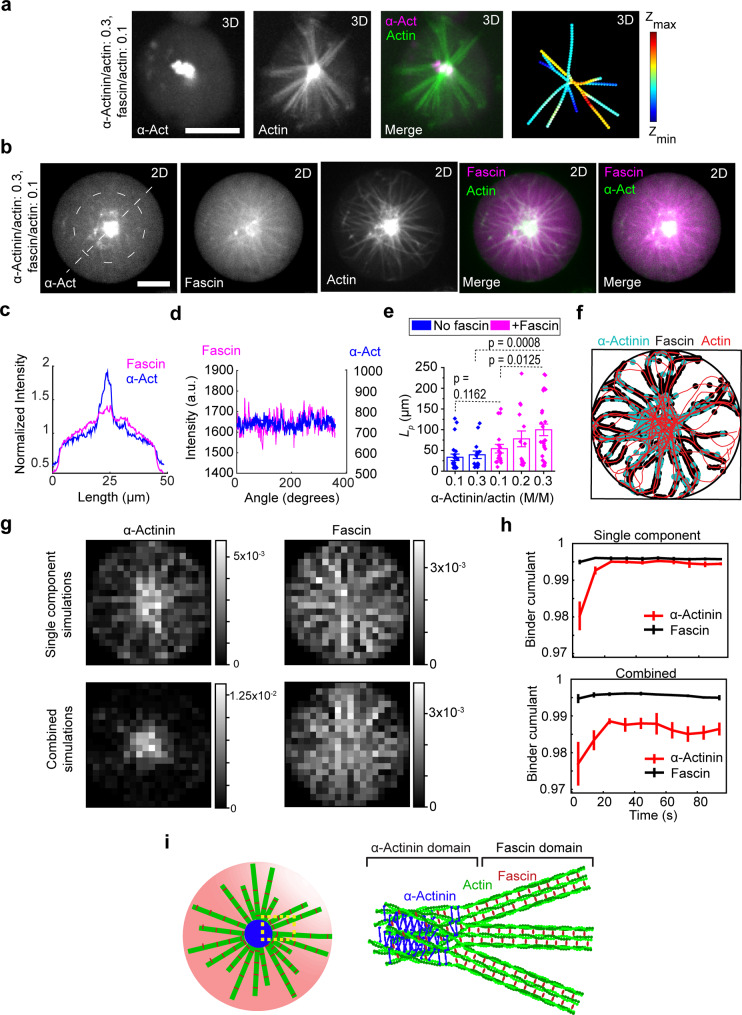
The proteins that make up the actin cytoskeleton can self-assemble into a variety of structures. In vitro experiments and coarse-grained simulations have shown that the actin crosslinking proteins α-actinin and fascin segregate into distinct domains in single actin bundles with a molecular size-dependent competition-based mechanism. Here, by encapsulating actin, α-actinin, and fascin in giant unilamellar vesicles (GUVs), we show that physical confinement can cause these proteins to form much more complex structures, including rings and asters at GUV peripheries and centers; the prevalence of different structures depends on GUV size. Strikingly, we found that α-actinin and fascin self-sort into separate domains in the aster structures with actin bundles whose apparent stiffness depends on the ratio of the relative concentrations of α-actinin and fascin. The observed boundary-imposed effect on protein sorting may be a general mechanism for creating emergent structures in biopolymer networks with multiple crosslinkers.
© 2021. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
