

SALM4 negatively regulates NMDA receptor function and fear memory consolidation
- PMID: 34588597
- PMCID: PMC8481232
- DOI: 10.1038/s42003-021-02656-3
SALM4 negatively regulates NMDA receptor function and fear memory consolidation
Abstract
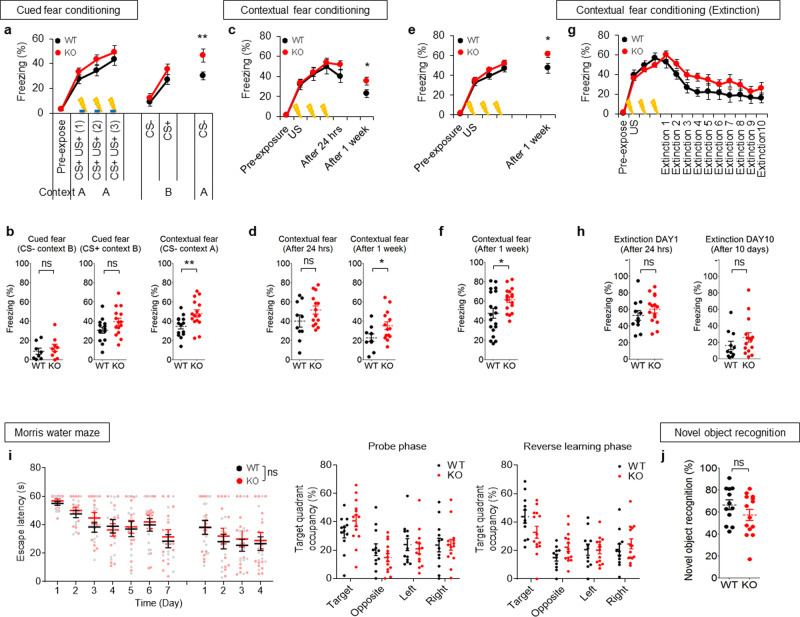
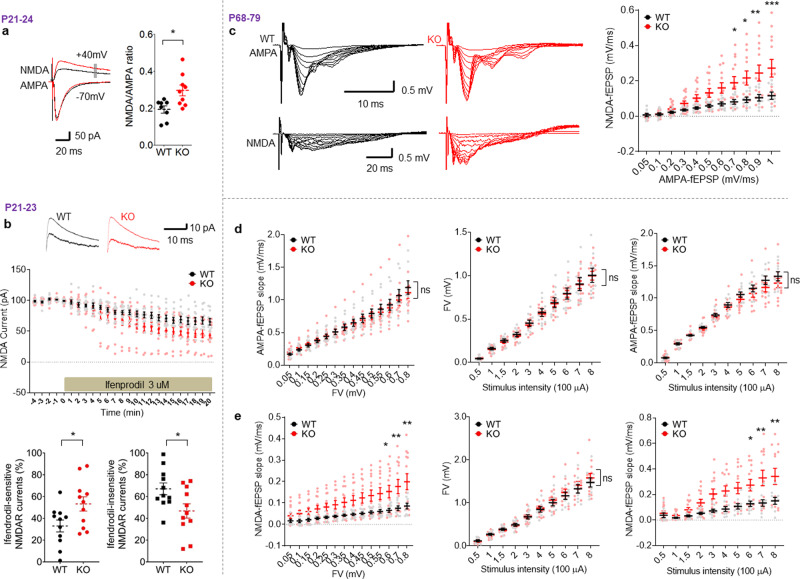
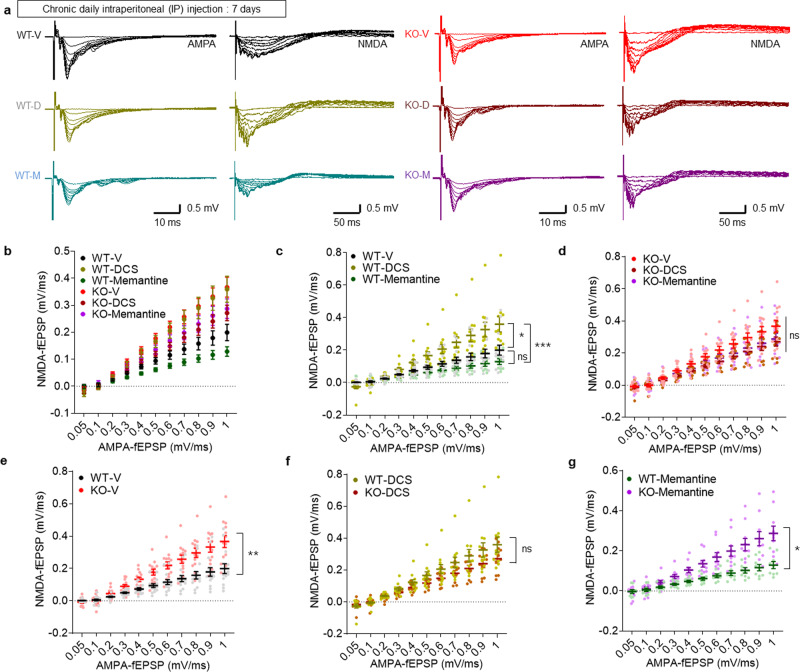
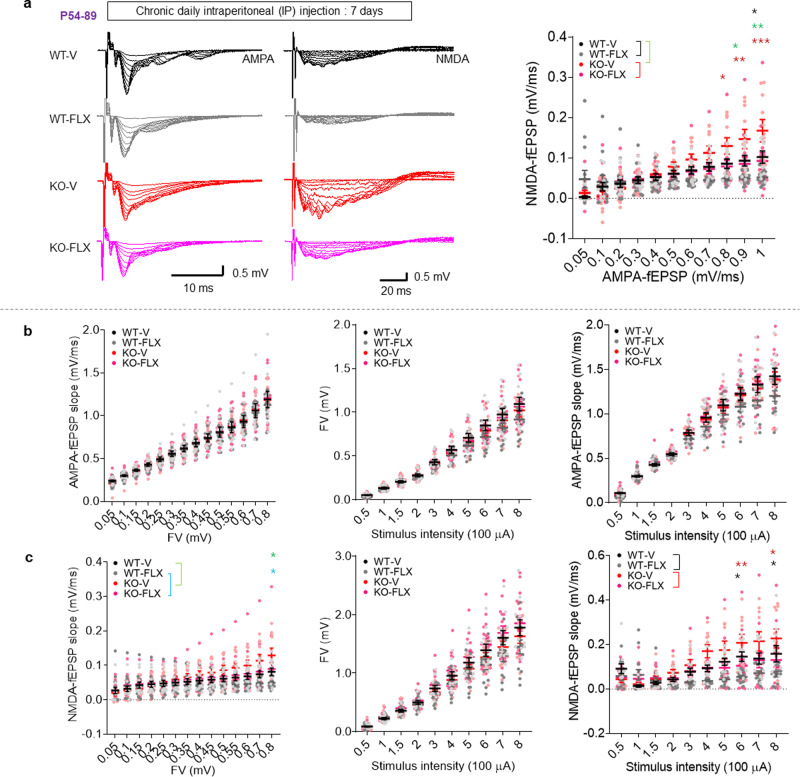
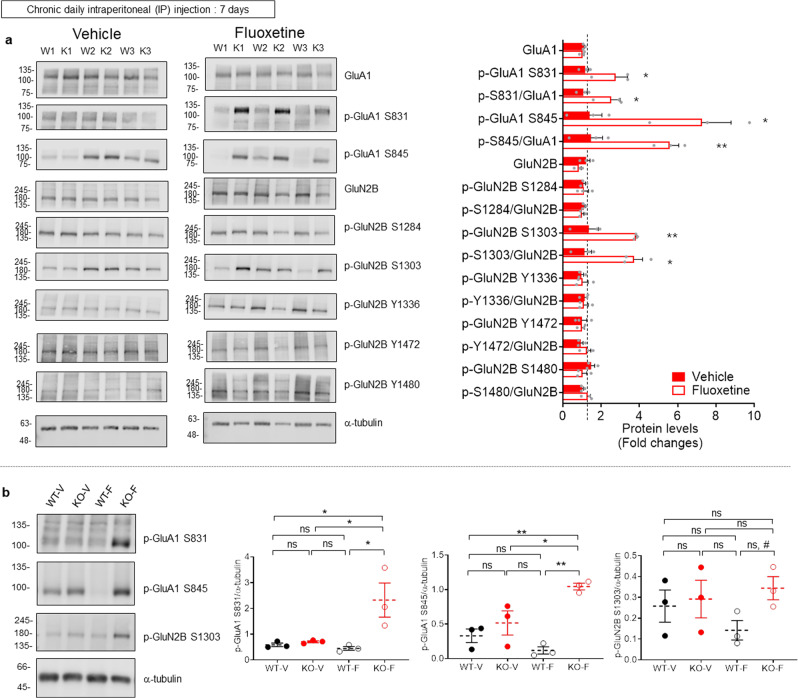
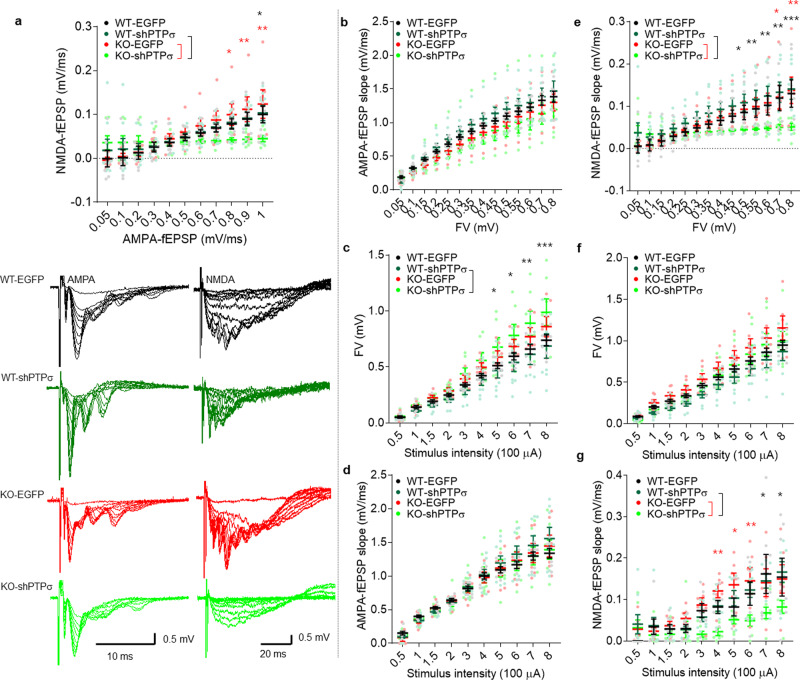
Many synaptic adhesion molecules positively regulate synapse development and function, but relatively little is known about negative regulation. SALM4/Lrfn3 (synaptic adhesion-like molecule 4/leucine rich repeat and fibronectin type III domain containing 3) inhibits synapse development by suppressing other SALM family proteins, but whether SALM4 also inhibits synaptic function and specific behaviors remains unclear. Here we show that SALM4-knockout (Lrfn3-/-) male mice display enhanced contextual fear memory consolidation (7-day post-training) but not acquisition or 1-day retention, and exhibit normal cued fear, spatial, and object-recognition memory. The Lrfn3-/- hippocampus show increased currents of GluN2B-containing N-methyl-D-aspartate (NMDA) receptors (GluN2B-NMDARs), but not α-amino-3-hydroxy-5-methyl-4-isoxazole propionate (AMPA) receptors (AMPARs), which requires the presynaptic receptor tyrosine phosphatase PTPσ. Chronic treatment of Lrfn3-/- mice with fluoxetine, a selective serotonin reuptake inhibitor used to treat excessive fear memory that directly inhibits GluN2B-NMDARs, normalizes NMDAR function and contextual fear memory consolidation in Lrfn3-/- mice, although the GluN2B-specific NMDAR antagonist ifenprodil was not sufficient to reverse the enhanced fear memory consolidation. These results suggest that SALM4 suppresses excessive GluN2B-NMDAR (not AMPAR) function and fear memory consolidation (not acquisition).
© 2021. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures



References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
