

Recent advances in cellular biosensor technology to investigate tau oligomerization
- PMID: 34589603
- PMCID: PMC8459642
- DOI: 10.1002/btm2.10231
Recent advances in cellular biosensor technology to investigate tau oligomerization
Abstract
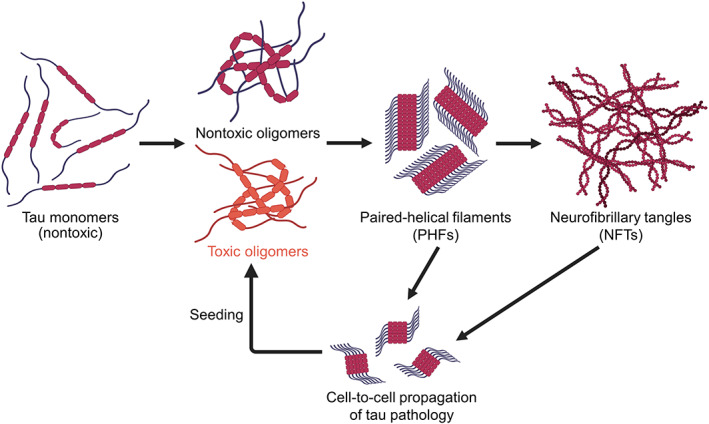
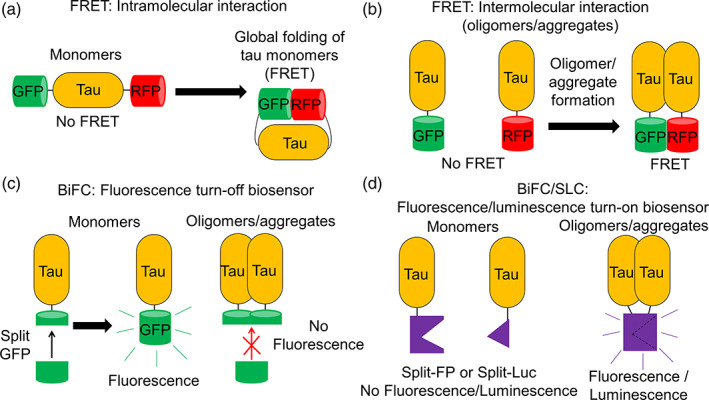
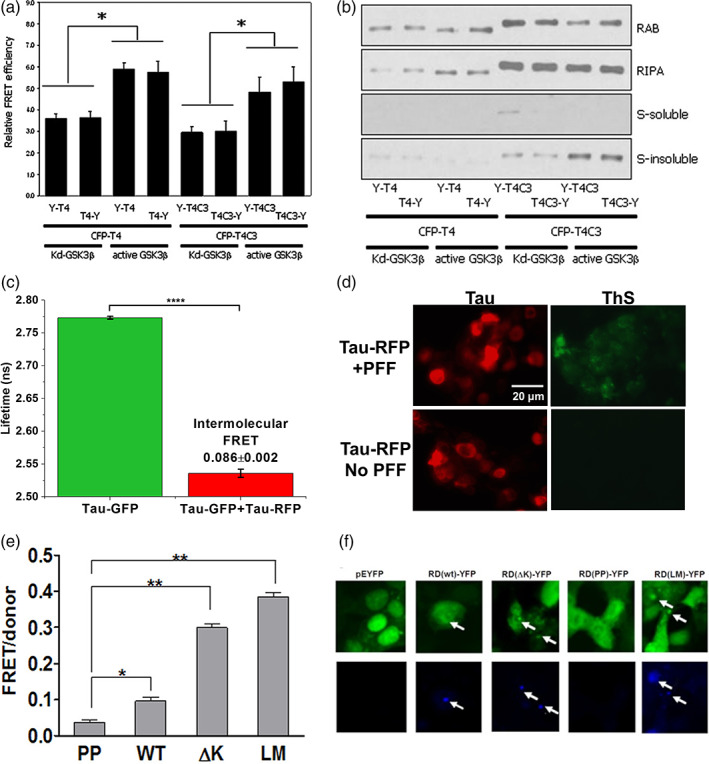
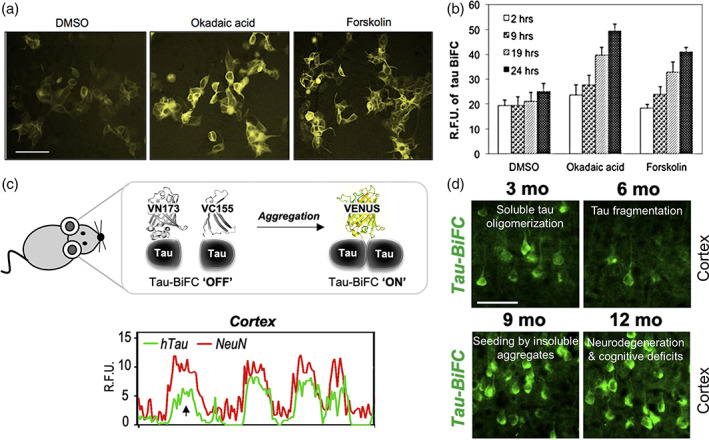
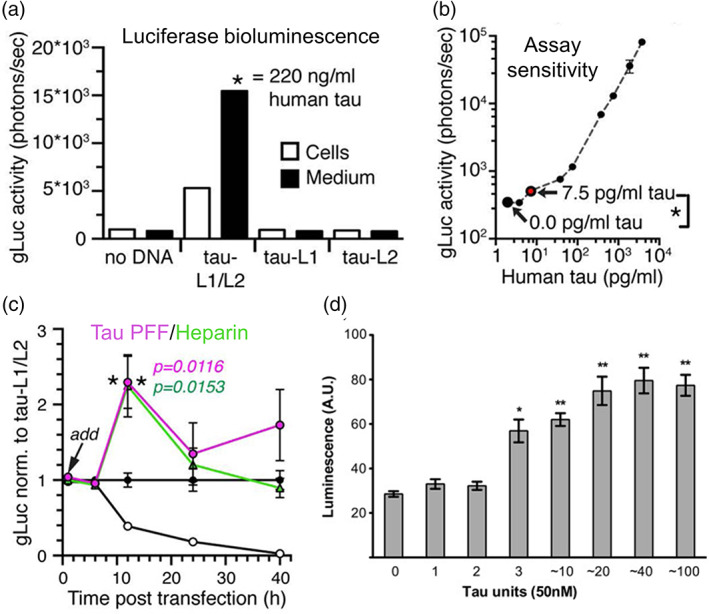
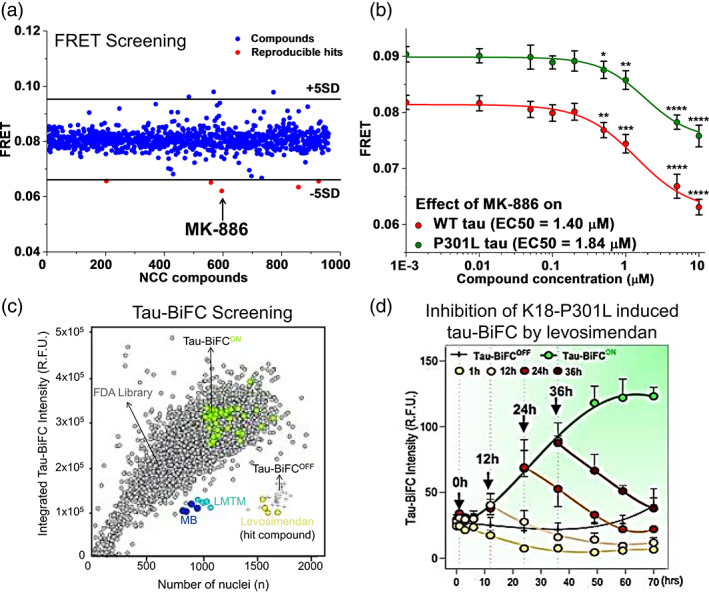
Tau is a microtubule binding protein which plays an important role in physiological functions but it is also involved in the pathogenesis of Alzheimer's disease and related tauopathies. While insoluble and β-sheet containing tau neurofibrillary tangles have been the histopathological hallmark of these diseases, recent studies suggest that soluble tau oligomers, which are formed prior to fibrils, are the primary toxic species. Substantial efforts have been made to generate tau oligomers using purified recombinant protein strategies to study oligomer conformations as well as their toxicity. However, no specific toxic tau species has been identified to date, potentially due to the lack of cellular environment. Hence, there is a need for cell-based models for direct monitoring of tau oligomerization and aggregation. This review will summarize the recent advances in the cellular biosensor technology, with a focus on fluorescence resonance energy transfer, bimolecular fluorescence complementation, and split luciferase complementation approaches, to monitor formation of tau oligomers and aggregates in living cells. We will discuss the applications of the cellular biosensors in examining the heterogeneous tau conformational ensembles and factors affecting tau self-assembly, as well as detecting cell-to-cell propagation of tau pathology. We will also compare the advantages and limitations of each type of tau biosensors, and highlight their translational applications in biomarker development and therapeutic discovery.
Keywords: Alzheimer's disease (AD); bimolecular fluorescence complementation (BiFC); cell‐based biosensor; conformational ensembles; fluorescence resonance energy transfer (FRET); high‐throughput screening (HTS); protein–protein interaction (PPI); split fluorescent protein complementation; split luciferase complementation (SLC); tau oligomerization.
© 2021 The Author. Bioengineering & Translational Medicine published by Wiley Periodicals LLC on behalf of American Institute of Chemical Engineers.
Conflict of interest statement
The author declares no conflict of interest.
Figures

References
-
- National Institute on Aging, NIH . Alzheimer's Disease Fact Sheet. 2021.
-
- Ballatore C, Lee VMY, Trojanowski JQ. Tau‐mediated neurodegeneration in Alzheimer's disease and related disorders. Nat Rev Neurosci. 2007;8(9):663‐672. - PubMed
-
- Avila J, Lucas JJ, Pérez M, Hernández F. Role of tau protein in both physiological and pathological conditions. Physiol Rev. 2004;84(2):361‐384. - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
