

A protein network map of head and neck cancer reveals PIK3CA mutant drug sensitivity
- PMID: 34591642
- PMCID: PMC9005332
- DOI: 10.1126/science.abf2911
A protein network map of head and neck cancer reveals PIK3CA mutant drug sensitivity
Abstract
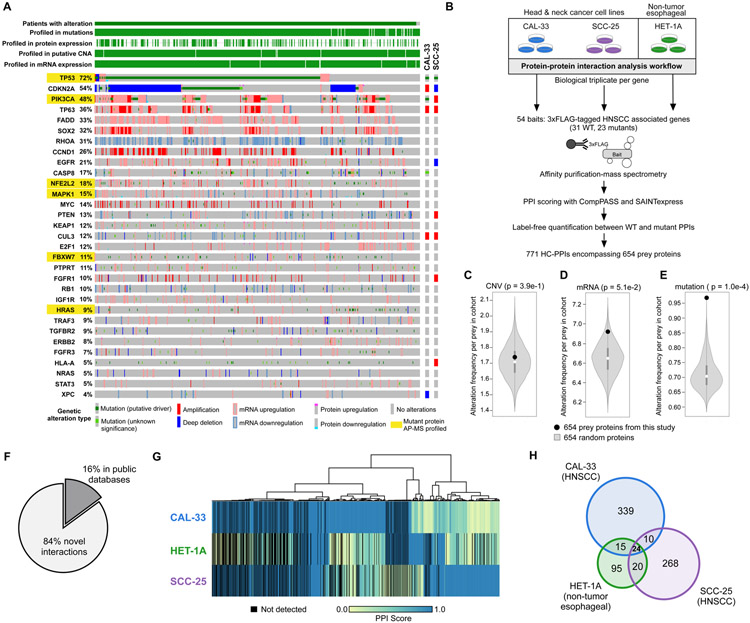
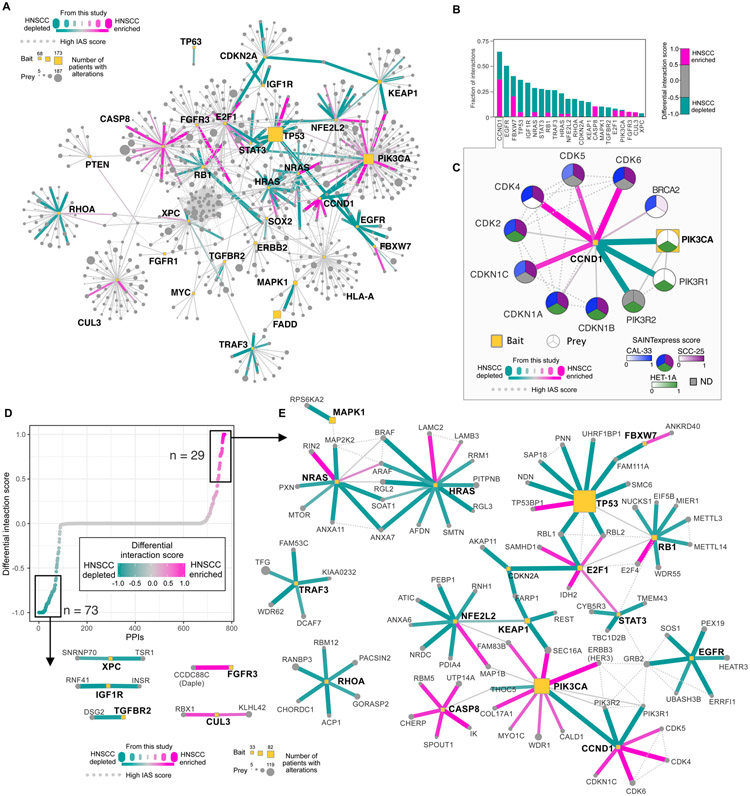
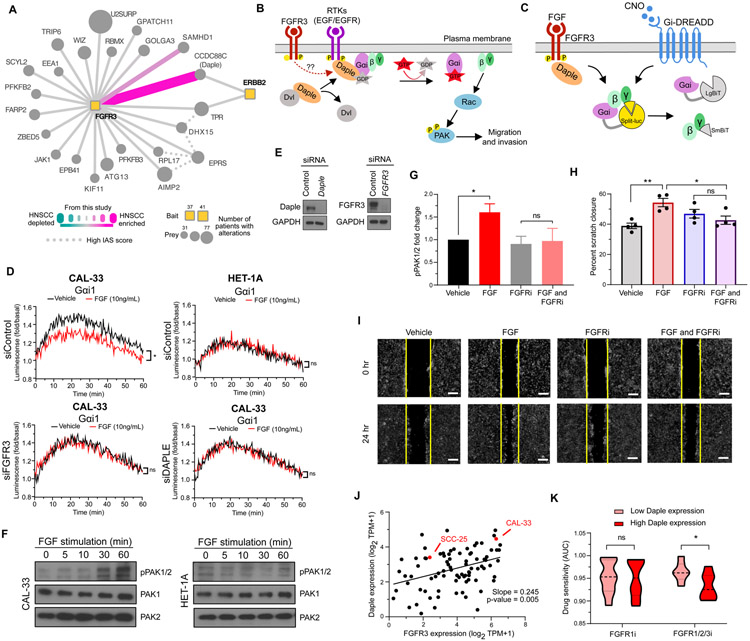
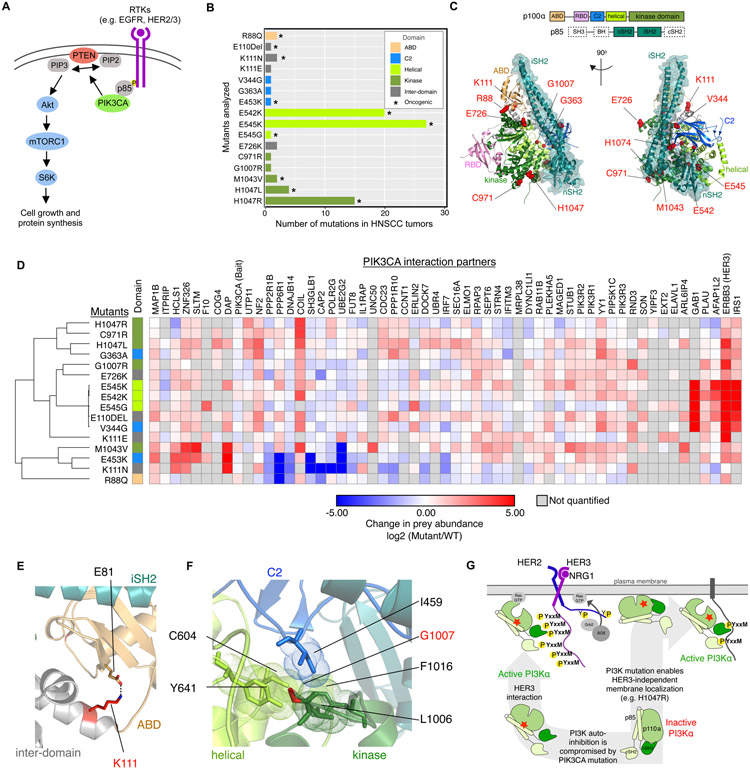
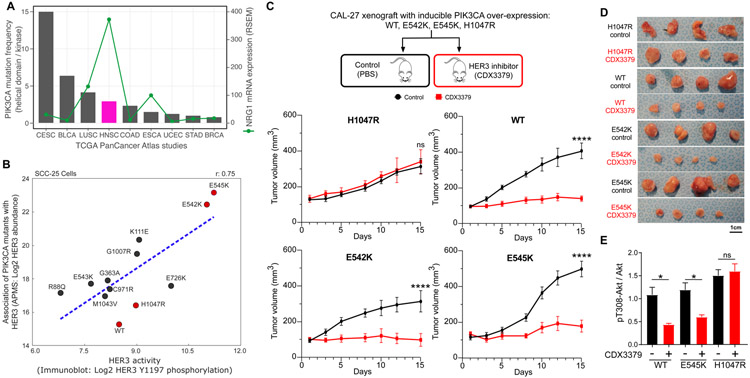
We outline a framework for elucidating tumor genetic complexity through multidimensional protein-protein interaction maps and apply it to enhancing our understanding of head and neck squamous cell carcinoma. This network uncovers 771 interactions from cancer and noncancerous cell states, including WT and mutant protein isoforms. Prioritization of cancer-enriched interactions reveals a previously unidentified association of the fibroblast growth factor receptor tyrosine kinase 3 with Daple, a guanine-nucleotide exchange factor, resulting in activation of Gαi- and p21-activated protein kinase 1/2 to promote cancer cell migration. Additionally, we observe mutation-enriched interactions between the human epidermal growth factor receptor 3 (HER3) receptor tyrosine kinase and PIK3CA (the alpha catalytic subunit of phosphatidylinositol 3-kinase) that can inform the response to HER3 inhibition in vivo. We anticipate that the application of this framework will be valuable for translating genetic alterations into a molecular and clinical understanding of the underlying biology of many disease areas.
Figures
Comment in
-
Identifying cancer drivers.Science. 2021 Oct;374(6563):38-39. doi: 10.1126/science.abl9080. Epub 2021 Sep 30. Science. 2021. PMID: 34591644
References
-
- Robinson D, Van Allen EM, Wu YM, Schultz N, Lonigro RJ, Mosquera JM, Montgomery B, Taplin ME, Pritchard CC, Attard G, Beltran H, Abida W, Bradley RK, Vinson J, Cao X, Vats P, Kunju LP, Hussain M, Feng FY, Tomlins SA, Cooney KA, Smith DC, Brennan C, Siddiqui J, Mehra R, Chen Y, Rathkopf DE, Morris MJ, Solomon SB, Durack JC, Reuter VE, Gopalan A, Gao J, Loda M, Lis RT, Bowden M, Balk SP, Gaviola G, Sougnez C, Gupta M, Yu EY, Mostaghel EA, Cheng HH, Mulcahy H, True LD, Plymate SR, Dvinge H, Ferraldeschi R, Flohr P, Miranda S, Zafeiriou Z, Tunariu N, Mateo J, Perez-Lopez R, Demichelis F, Robinson BD, Sboner A, Schiffman M, Nanus DM, Tagawa ST, Sigaras A, Eng KW, Elemento O, Sboner A, Heath EI, Scher HI, Pienta KJ, Kantoff P, de Bono JS, Rubin MA, Nelson PS, Garraway LA, Sawyers CL, Chinnaiyan AM, Integrative Clinical Genomics of Advanced Prostate Cancer. Cell. 162, 454 (2015). - PubMed
-
- Biankin AV, Waddell N, Kassahn KS, Gingras MC, Muthuswamy LB, Johns AL, Miller DK, Wilson PJ, Patch AM, Wu J, Chang DK, Cowley MJ, Gardiner BB, Song S, Harliwong I, Idrisoglu S, Nourse C, Nourbakhsh E, Manning S, Wani S, Gongora M, Pajic M, Scarlett CJ, Gill AJ, Pinho AV, Rooman I, Anderson M, Holmes O, Leonard C, Taylor D, Wood S, Xu Q, Nones K, Fink JL, Christ A, Bruxner T, Cloonan N, Kolle G, Newell F, Pinese M, Mead RS, Humphris JL, Kaplan W, Jones MD, Colvin EK, Nagrial AM, Humphrey ES, Chou A, Chin VT, Chantrill LA, Mawson A, Samra JS, Kench JG, Lovell JA, Daly RJ, Merrett ND, Toon C, Epari K, Nguyen NQ, Barbour A, Zeps N, Australian Pancreatic Cancer Genome, Initiative, Kakkar N, Zhao F, Wu YQ, Wang M, Muzny DM, Fisher WE, Brunicardi FC, Hodges SE, Reid JG, Drummond J, Chang K, Han Y, Lewis LR, Dinh H, Buhay CJ, Beck T, Timms L, Sam M, Begley K, Brown A, Pai D, Panchal A, Buchner N, De Borja R, Denroche RE, Yung CK, Serra S, Onetto N, Mukhopadhyay D, Tsao MS, Shaw PA, Petersen GM, Gallinger S, Hruban RH, Maitra A, Iacobuzio-Donahue CA, Schulick RD, Wolfgang CL, Morgan RA, Lawlor RT, Capelli P, Corbo V, Scardoni M, Tortora G, Tempero MA, Mann KM, Jenkins NA, Perez-Mancera PA, Adams DJ, Largaespada DA, Wessels LF, Rust AG, Stein LD, Tuveson DA, Copeland NG, Musgrove EA, Scarpa A, Eshleman JR, Hudson TJ, Sutherland RL, Wheeler DA, Pearson JV, McPherson JD, Gibbs RA, Grimmond SM, Pancreatic cancer genomes reveal aberrations in axon guidance pathway genes. Nature. 491, 399–405 (2012). - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 GM109176/GM/NIGMS NIH HHS/United States
- R50 CA243885/CA/NCI NIH HHS/United States
- P41 GM103504/GM/NIGMS NIH HHS/United States
- F32 CA239333/CA/NCI NIH HHS/United States
- R01 DE026644/DE/NIDCR NIH HHS/United States
- R01 DE028289/DE/NIDCR NIH HHS/United States
- R35 CA231998/CA/NCI NIH HHS/United States
- R01 DE023685/DE/NIDCR NIH HHS/United States
- U24 CA184427/CA/NCI NIH HHS/United States
- U01 DE028227/DE/NIDCR NIH HHS/United States
- R01 CA247551/CA/NCI NIH HHS/United States
- R01 DE026870/DE/NIDCR NIH HHS/United States
- U54 CA209891/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
