

Genetically targeting the BATF family transcription factors BATF and BATF3 in the mouse abrogates effector T cell activities and enables long-term heart allograft survival
- PMID: 34599765
- PMCID: PMC8813885
- DOI: 10.1111/ajt.16861
Genetically targeting the BATF family transcription factors BATF and BATF3 in the mouse abrogates effector T cell activities and enables long-term heart allograft survival
Abstract
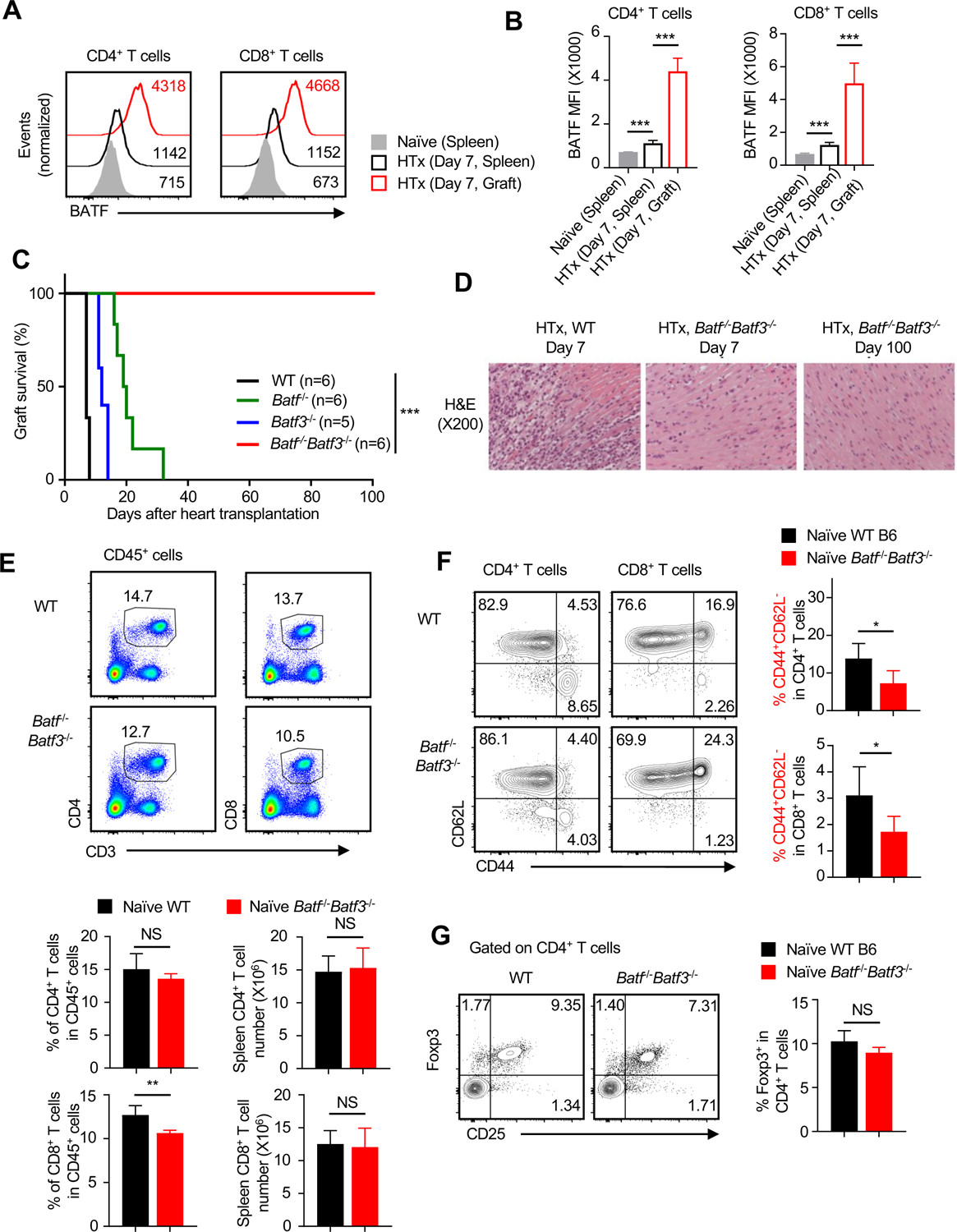
T cells must be activated and become effectors first before executing allograft rejection, a process that is regulated by diverse signals and transcription factors. In this study, we studied the basic leucine zipper ATF-like transcription factor (BATF) family members in regulating T cell activities in a heart transplant model and found that mice deficient for both BATF and BATF3 (Batf-/- Batf3-/- mice) spontaneously accept the heart allografts long-term without tolerizing therapies. Similarly, adoptive transfer of wild type T cells into Rag1-/- hosts induced prompt rejection of heart and skin allografts, whereas the Batf-/- Batf3-/- T cells failed to do so. Analyses of graft-infiltrating cells showed that Batf-/- Batf3-/- T cells infiltrate the graft but fail to acquire an effector phenotype (CD44high KLRG1+ ). Co-transfer experiments in a T cell receptor transgenic TEa model revealed that the Batf-/- Batf3-/- T cells fail to expand in vivo, retain a quiescent phenotype (CD62L+ CD127+ ), and unable to produce effector cytokines to alloantigen stimulation, which contrasted sharply to that of wild type T cells. Together, our data demonstrate that the BATF and BATF3 are critical regulators of T effector functions, thus making them attractive targets for therapeutic interventions in transplant settings.
Keywords: T cell biology; basic (laboratory) research/science; immunobiology; tolerance: experimental.
© 2021 The American Society of Transplantation and the American Society of Transplant Surgeons.
Conflict of interest statement
Disclosure
The authors of this manuscript have no conflicts of interest to disclose as described by the
Figures





References
-
- Murphy TL, Tussiwand R, Murphy KM. Specificity through cooperation: BATF-IRF interactions control immune-regulatory networks. Nat Rev Immunol. 2013;13(7):499–509. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
