

Molecular basis of a bacterial-amphibian symbiosis revealed by comparative genomics, modeling, and functional testing
- PMID: 34601502
- PMCID: PMC8857215
- DOI: 10.1038/s41396-021-01121-7
Molecular basis of a bacterial-amphibian symbiosis revealed by comparative genomics, modeling, and functional testing
Abstract
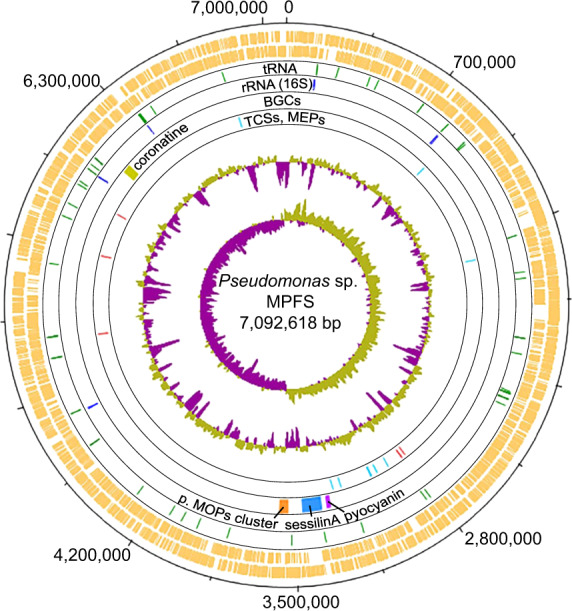
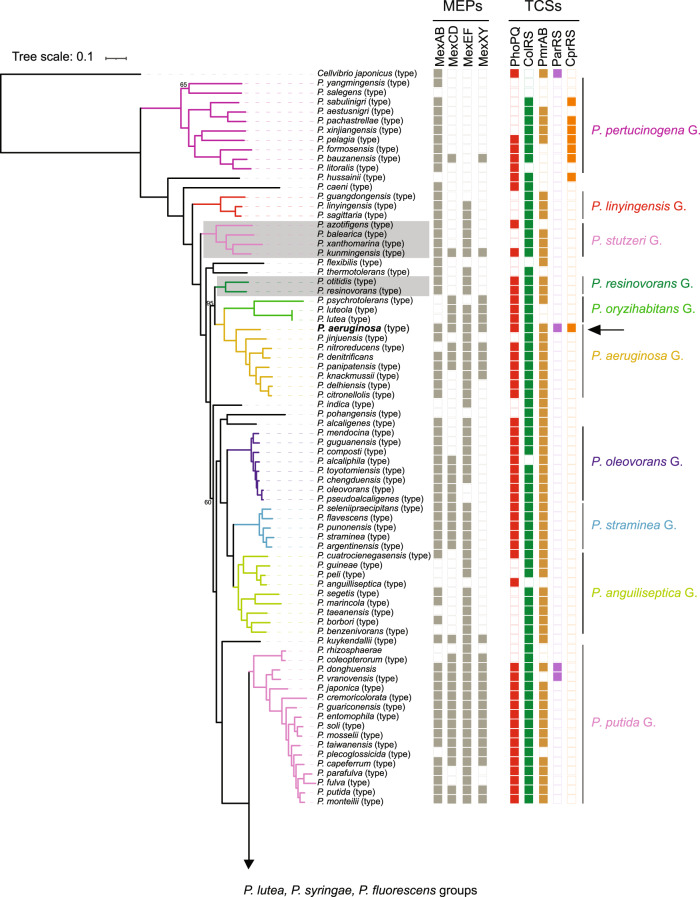
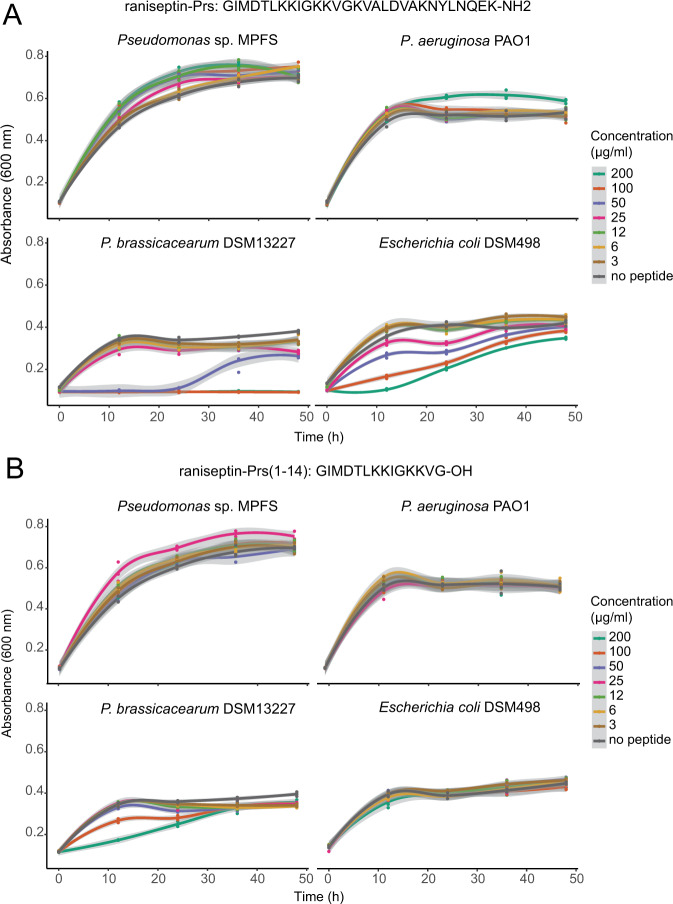
The molecular bases for the symbiosis of the amphibian skin microbiome with its host are poorly understood. Here, we used the odor-producer Pseudomonas sp. MPFS and the treefrog Boana prasina as a model to explore bacterial genome determinants and the resulting mechanisms facilitating symbiosis. Pseudomonas sp. MPFS and its closest relatives, within a new clade of the P. fluoresens Group, have large genomes and were isolated from fishes and plants, suggesting environmental plasticity. We annotated 16 biosynthetic gene clusters from the complete genome sequence of this strain, including those encoding the synthesis of compounds with known antifungal activity and of odorous methoxypyrazines that likely mediate sexual interactions in Boana prasina. Comparative genomics of Pseudomonas also revealed that Pseudomonas sp. MPFS and its closest relatives have acquired specific resistance mechanisms against host antimicrobial peptides (AMPs), specifically two extra copies of a multidrug efflux pump and the same two-component regulatory systems known to trigger adaptive resistance to AMPs in P. aeruginosa. Subsequent molecular modeling indicated that these regulatory systems interact with an AMP identified in Boana prasina through the highly acidic surfaces of the proteins comprising their sensory domains. In agreement with a symbiotic relationship and a highly selective antibacterial function, this AMP did not inhibit the growth of Pseudomonas sp. MPFS but inhibited the growth of another Pseudomonas species and Escherichia coli in laboratory tests. This study provides deeper insights into the molecular interaction of the bacteria-amphibian symbiosis and highlights the role of specific adaptive resistance toward AMPs of the hosts.
© 2021. The Author(s), under exclusive licence to International Society for Microbial Ecology.
Conflict of interest statement
The authors declare no competing interests.
Figures




References
-
- Toledo RC, Jared C. Cutaneous granular glands and amphibian venoms. Comp Biochem Physiol A Physiol. 1995;111:1–29.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
