

Distinct B cell subsets in Peyer's patches convey probiotic effects by Limosilactobacillus reuteri
- PMID: 34602091
- PMCID: PMC8487498
- DOI: 10.1186/s40168-021-01128-4
Distinct B cell subsets in Peyer's patches convey probiotic effects by Limosilactobacillus reuteri
Abstract
Background: Intestinal Peyer's patches (PPs) form unique niches for bacteria-immune cell interactions that direct host immunity and shape the microbiome. Here we investigate how peroral administration of probiotic bacterium Limosilactobacillus reuteri R2LC affects B lymphocytes and IgA induction in the PPs, as well as the downstream consequences on intestinal microbiota and susceptibility to inflammation.
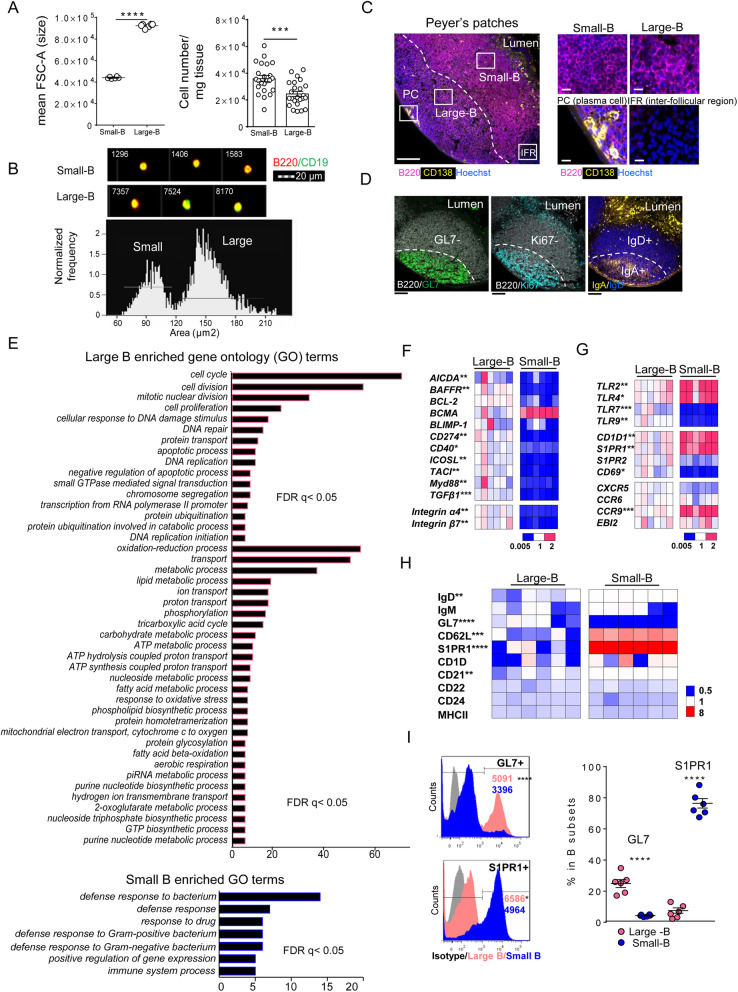
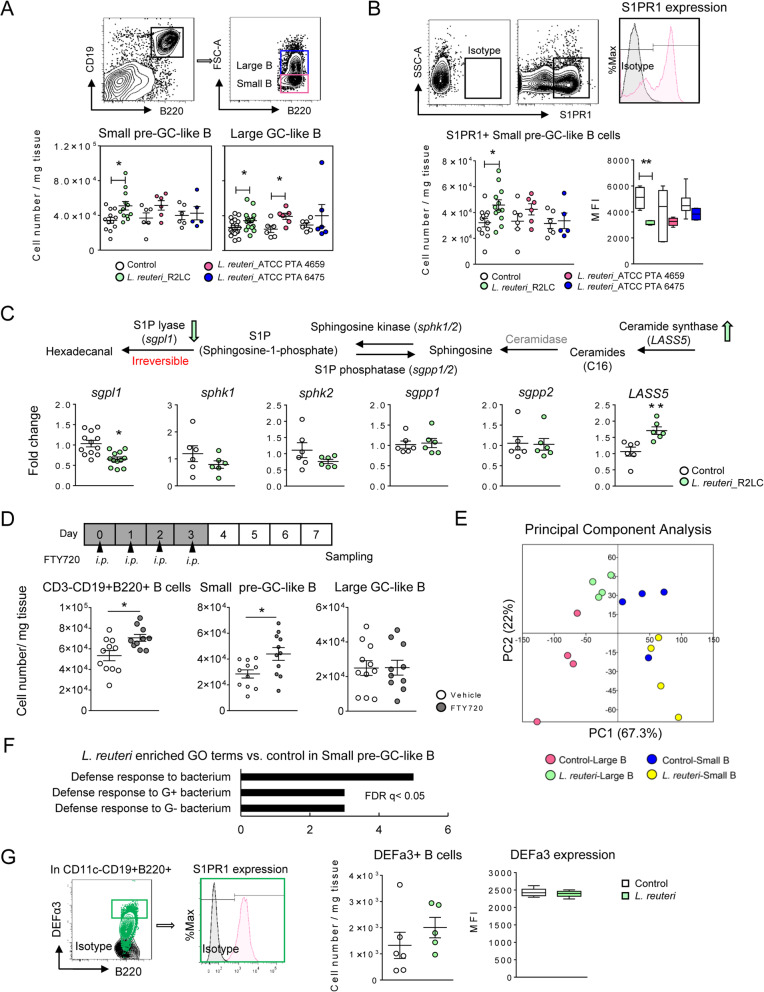
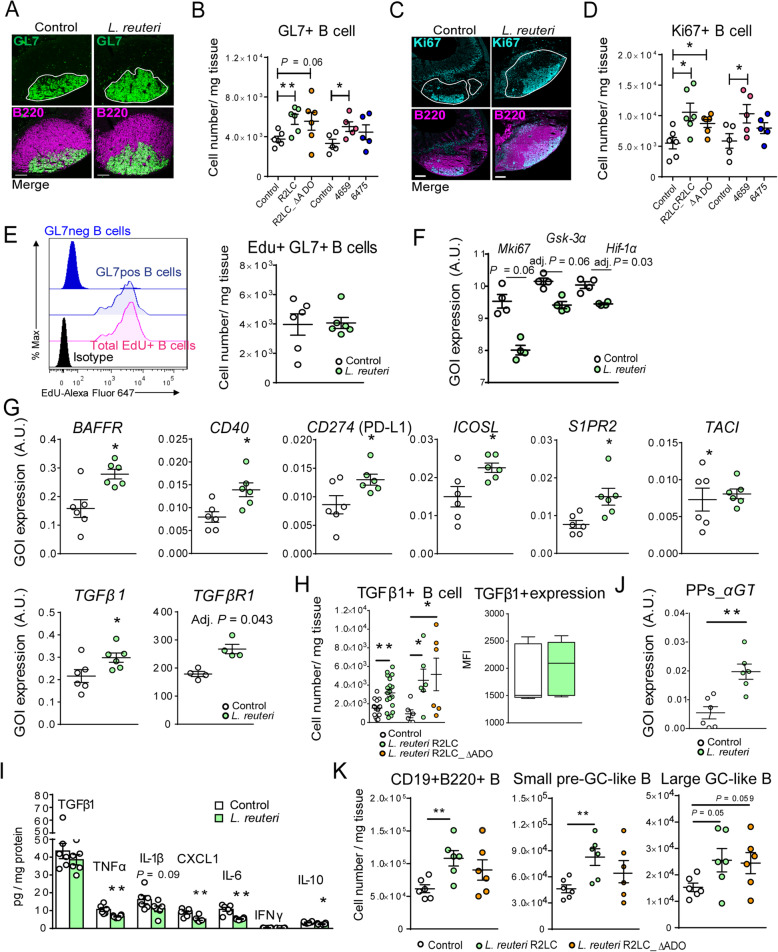
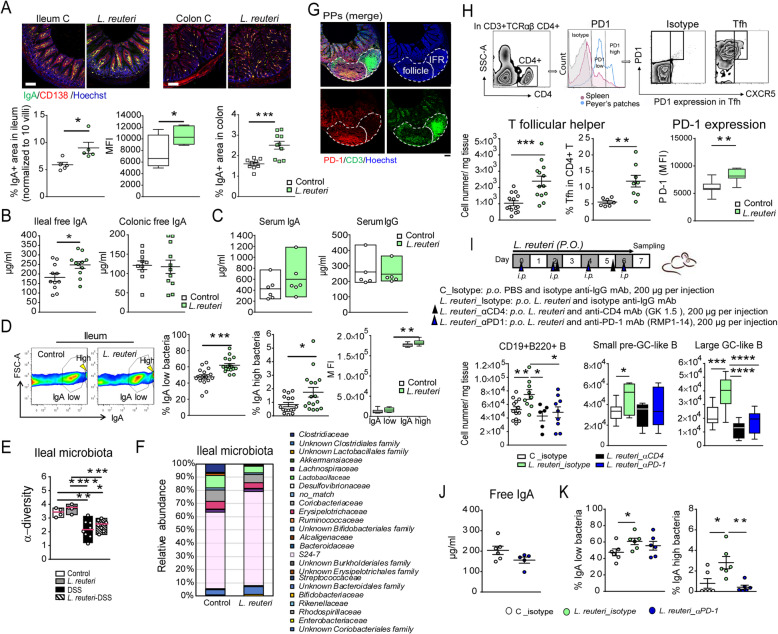
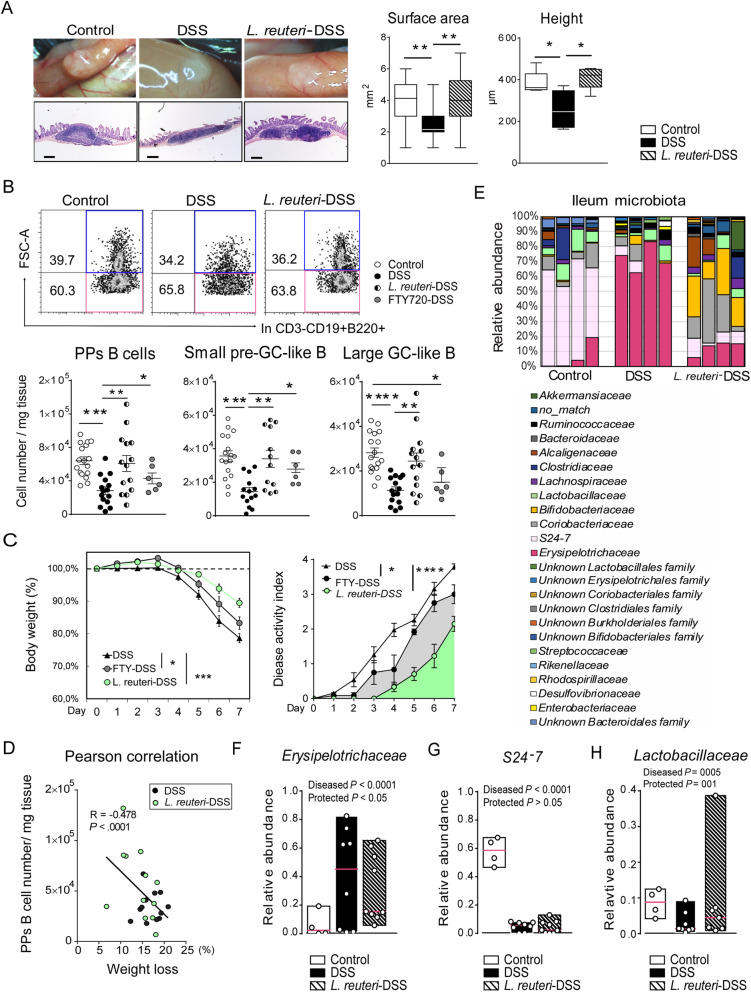
Results: The B cells of PPs were separated by size to circumvent activation-dependent cell identification biases due to dynamic expression of markers, which resulted in two phenotypically, transcriptionally, and spatially distinct subsets: small IgD+/GL7-/S1PR1+/Bcl6, CCR6-expressing pre-germinal center (GC)-like B cells with innate-like functions located subepithelially, and large GL7+/S1PR1-/Ki67+/Bcl6, CD69-expressing B cells with strong metabolic activity found in the GC. Peroral L. reuteri administration expanded both B cell subsets and enhanced the innate-like properties of pre-GC-like B cells while retaining them in the sub-epithelial compartment by increased sphingosine-1-phosphate/S1PR1 signaling. Furthermore, L. reuteri promoted GC-like B cell differentiation, which involved expansion of the GC area and autocrine TGFβ-1 activation. Consequently, PD-1-T follicular helper cell-dependent IgA induction and production was increased by L. reuteri, which shifted the intestinal microbiome and protected against dextran-sulfate-sodium induced colitis and dysbiosis.
Conclusions: The Peyer's patches sense, enhance and transmit probiotic signals by increasing the numbers and effector functions of distinct B cell subsets, resulting in increased IgA production, altered intestinal microbiota, and protection against inflammation. Video abstract.
Keywords: Gut microbiome; Inflammatory bowel disease; Innate-like B lymphocytes; PD-1 dependent; Probiotics; R2LC.
© 2021. The Author(s).
Conflict of interest statement
The authors report no potential conflict of interest.
Figures
References
-
- Zheng J, Wittouck S, Salvetti E, Franz CMAP, Harris HMB, Mattarelli P, O’Toole PW, Pot B, Vandamme P, Walter J, Watanabe K, Wuyts S, Felis GE, Gänzle MG, Lebeer S. A taxonomic note on the genus Lactobacillus: description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int J Syst Evol Microbiol. 2020;70(4):2782–2858. doi: 10.1099/ijsem.0.004107. - DOI - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
