

TRF1 Depletion Reveals Mutual Regulation Between Telomeres, Kinetochores, and Inner Centromeres in Mouse Oocytes
- PMID: 34604243
- PMCID: PMC8486315
- DOI: 10.3389/fcell.2021.749116
TRF1 Depletion Reveals Mutual Regulation Between Telomeres, Kinetochores, and Inner Centromeres in Mouse Oocytes
Abstract
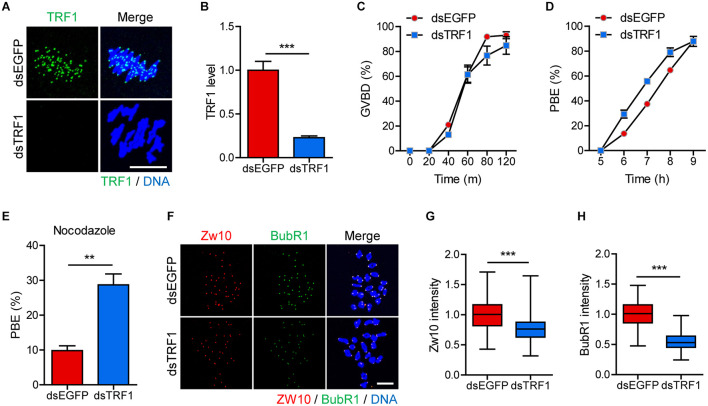
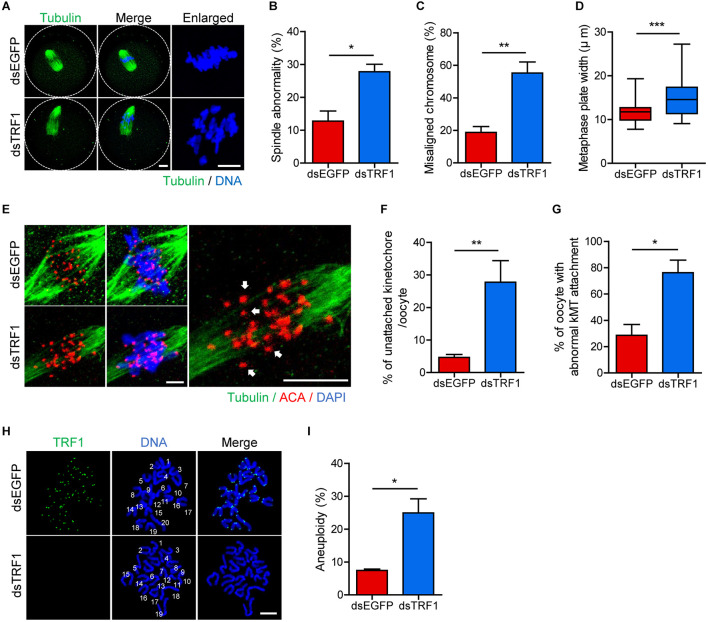
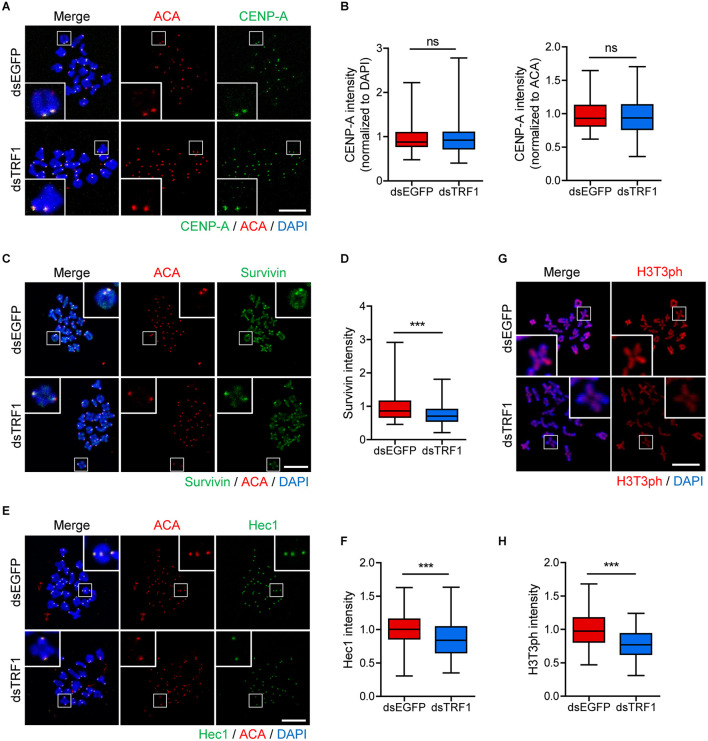
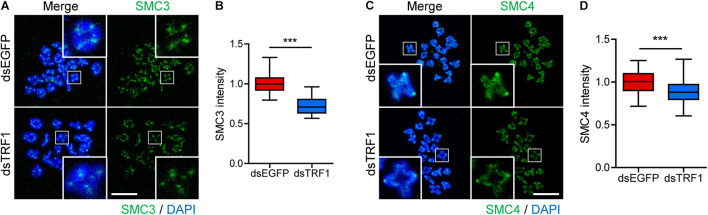
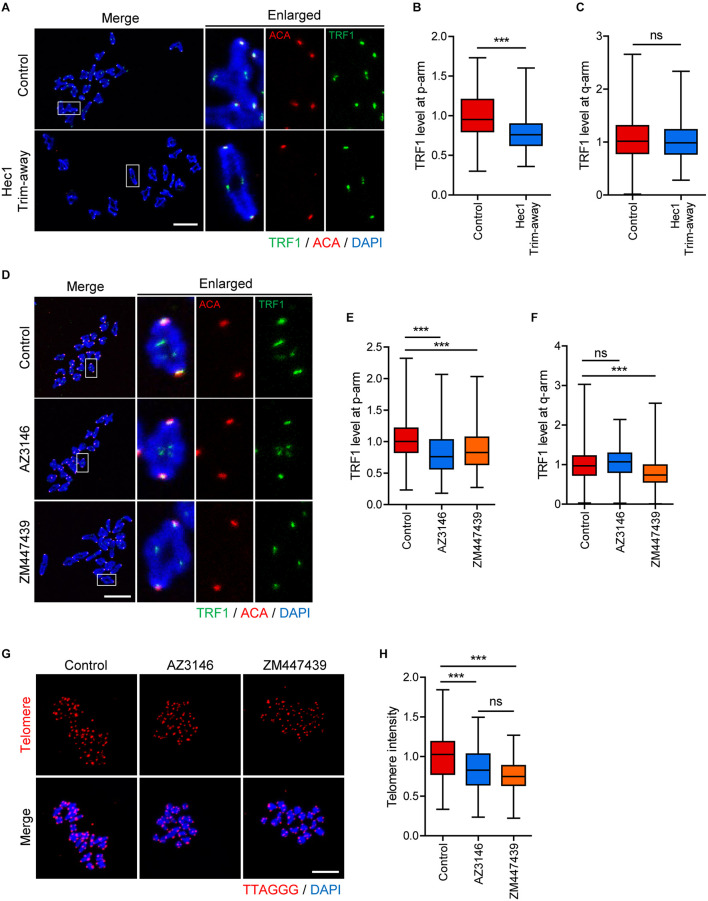
In eukaryotic chromosomes, the centromere and telomere are two specialized structures that are essential for chromosome stability and segregation. Although centromeres and telomeres often are located in close proximity to form telocentric chromosomes in mice, it remained unclear whether these two structures influence each other. Here we show that TRF1 is required for inner centromere and kinetochore assembly in addition to its role in telomere protection in mouse oocytes. TRF1 depletion caused premature chromosome segregation by abrogating the spindle assembly checkpoint (SAC) and impairing kinetochore-microtubule (kMT) attachment, which increased the incidence of aneuploidy. Notably, TRF1 depletion disturbed the localization of Survivin and Ndc80/Hec1 at inner centromeres and kinetochores, respectively. Moreover, SMC3 and SMC4 levels significantly decreased after TRF1 depletion, suggesting that TRF1 is involved in chromosome cohesion and condensation. Importantly, inhibition of inner centromere or kinetochore function led to a significant decrease in TRF1 level and telomere shortening. Therefore, our results suggest that telomere integrity is required to preserve inner centromere and kinetochore architectures, and vice versa, suggesting mutual regulation between telomeres and centromeres.
Keywords: TRF1; centromere; kinetochore; oocyte; telomere.
Copyright © 2021 Jeon and Oh.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
LinkOut - more resources
Full Text Sources
Miscellaneous