

A Modular Biomaterial Scaffold-Based Vaccine Elicits Durable Adaptive Immunity to Subunit SARS-CoV-2 Antigens
- PMID: 34605223
- PMCID: PMC8652677
- DOI: 10.1002/adhm.202101370
A Modular Biomaterial Scaffold-Based Vaccine Elicits Durable Adaptive Immunity to Subunit SARS-CoV-2 Antigens
Abstract
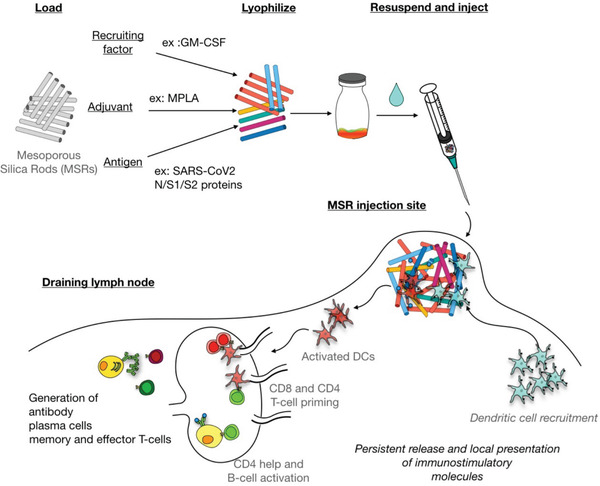
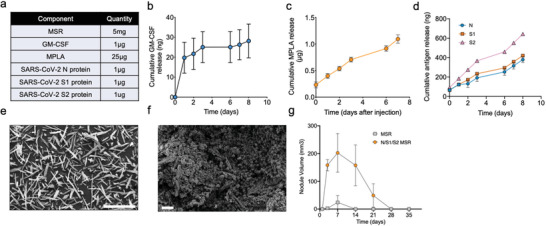
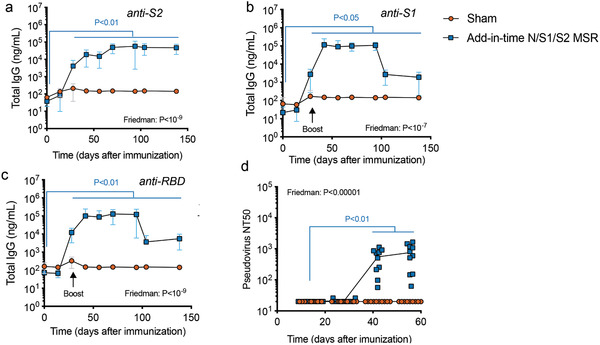
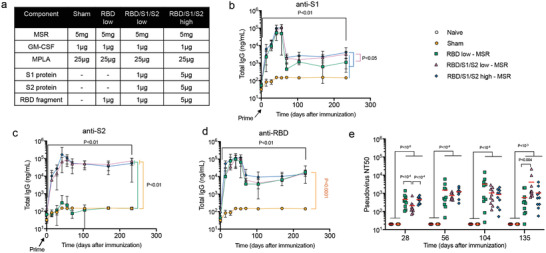
The coronavirus disease 2019 (COVID-19) pandemic demonstrates the importance of generating safe and efficacious vaccines that can be rapidly deployed against emerging pathogens. Subunit vaccines are considered among the safest, but proteins used in these typically lack strong immunogenicity, leading to poor immune responses. Here, a biomaterial COVID-19 vaccine based on a mesoporous silica rods (MSRs) platform is described. MSRs loaded with granulocyte-macrophage colony-stimulating factor (GM-CSF), the toll-like receptor 4 (TLR-4) agonist monophosphoryl lipid A (MPLA), and SARS-CoV-2 viral protein antigens slowly release their cargo and form subcutaneous scaffolds that locally recruit and activate antigen-presenting cells (APCs) for the generation of adaptive immunity. MSR-based vaccines generate robust and durable cellular and humoral responses against SARS-CoV-2 antigens, including the poorly immunogenic receptor binding domain (RBD) of the spike (S) protein. Persistent antibodies over the course of 8 months are found in all vaccine configurations tested and robust in vitro viral neutralization is observed both in a prime-boost and a single-dose regimen. These vaccines can be fully formulated ahead of time or stored lyophilized and reconstituted with an antigen mixture moments before injection, which can facilitate its rapid deployment against emerging SARS-CoV-2 variants or new pathogens. Together, the data show a promising COVID-19 vaccine candidate and a generally adaptable vaccine platform against infectious pathogens.
Keywords: COVID-19; SARS-CoV-2; antibodies; cytotoxic T-cells; mesoporous silica rods; monophosphoryl lipid A (MPLA); recombinant proteins; vaccines.
© 2021 Wiley-VCH GmbH.
Conflict of interest statement
Several authors are inventors on patent applications related to this technology (E.D, D.J.M); Novartis, sponsored research (D.J.M); Immulus, equity (D.J.M.), Attivare Therapeutics, equity (E.D., D.J.M., F.L., B.S.). David Walt has a financial interest in Quanterix Corporation, a company that develops an ultra‐sensitive digital immunoassay platform. He is an inventor of the Simoa technology, a founder of the company and also serves on its Board of Directors. Dr. Walt's interests were reviewed and are managed by BWH and Partners Healthcare in accordance with their conflict‐of‐interest policies.
Figures


References
-
- World Health Organization. WHO Coronavirus (COVID‐19) Dashboard. Available at: https://covid19.who.int/
-
- Gupta R. K., Harrison E. M., Ho A., Docherty A. B., Knight S. R., Van Smeden M., Abubakar I., Lipman M., Quartagno M., Pius R., Buchan I., Carson G., Drake T. M., Dunning J., Fairfield C. J., Gamble C., Green C. A., Halpin S., Hardwick H. E., Holden K. A., Horby P. W., Jackson C., Mclean K. A., Merson L., Nguyen‐Van‐Tam J. S., Norman L., Olliaro P. L., Pritchard M. G., Russell C. D., Scott‐Brown J., et al. Lancet Respir. Med. 2021, 9, 349. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
