

Heme-Dependent Siderophore Utilization Promotes Iron-Restricted Growth of the Staphylococcus aureus hemB Small-Colony Variant
- PMID: 34606375
- PMCID: PMC8604074
- DOI: 10.1128/JB.00458-21
Heme-Dependent Siderophore Utilization Promotes Iron-Restricted Growth of the Staphylococcus aureus hemB Small-Colony Variant
Abstract
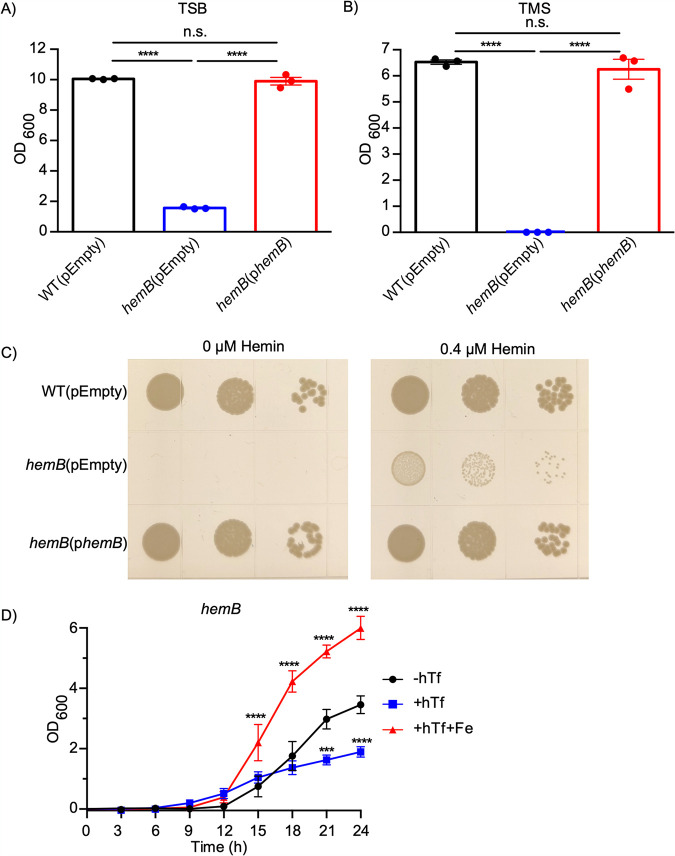
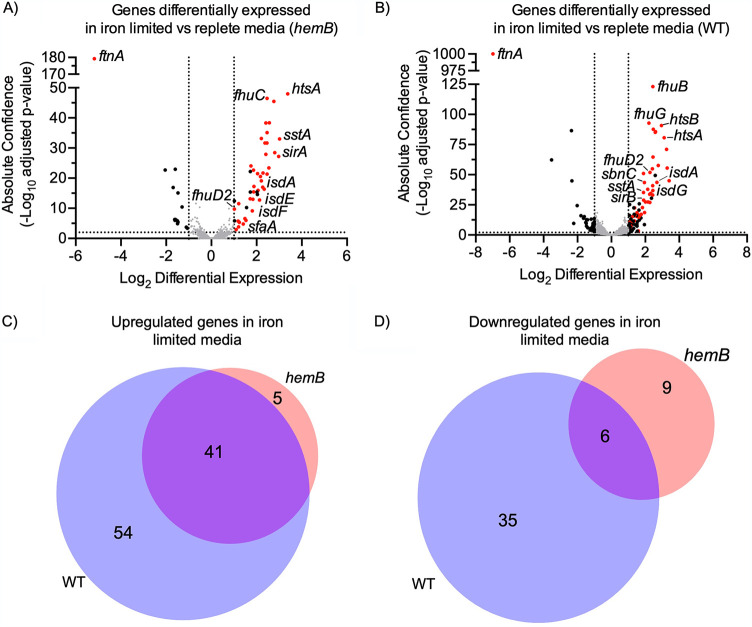
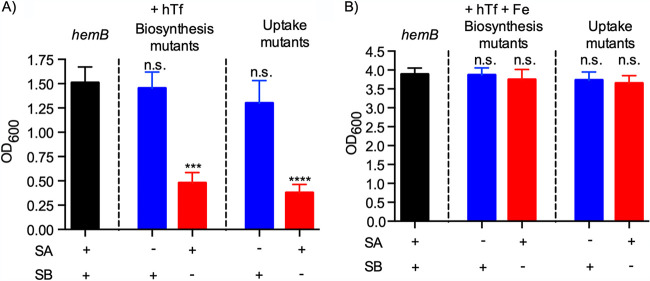
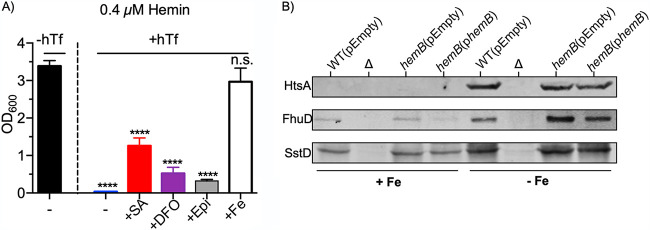
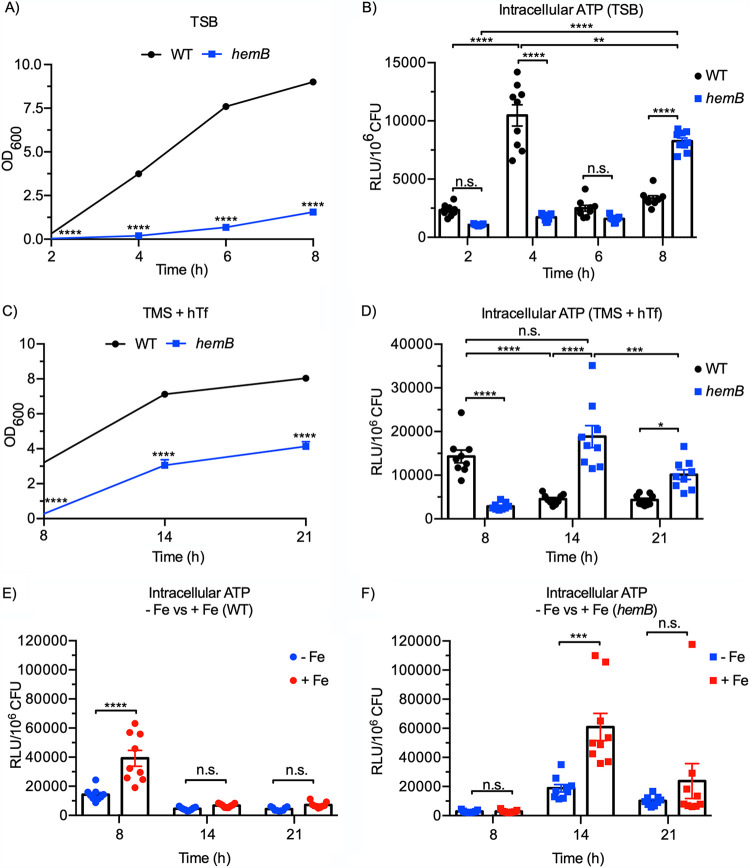
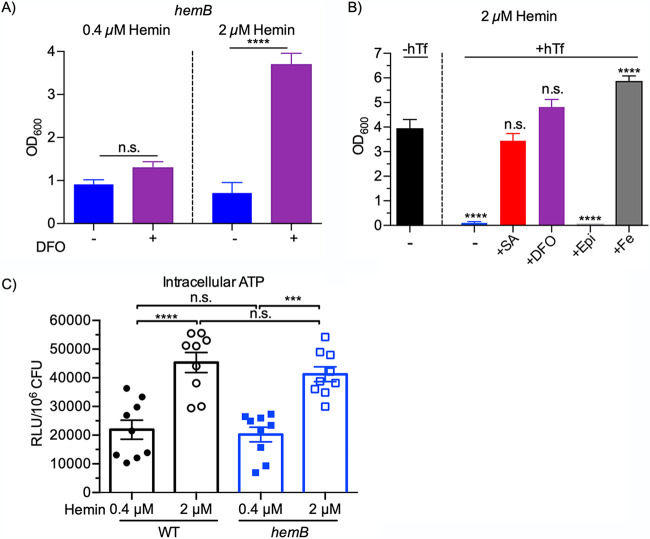
Respiration-deficient Staphylococcus aureus small-colony variants (SCVs) frequently cause persistent infections, which necessitates they acquire iron, yet how SCVs obtain iron remains unknown. To address this, we created a stable hemB mutant from S. aureus USA300 strain LAC. The hemB SCV utilized exogenously supplied hemin but was attenuated for growth under conditions of iron starvation. Transcriptome sequencing (RNA-seq) showed that both wild-type (WT) S. aureus and the hemB mutant sense and respond to iron starvation; however, growth assays show that the hemB mutant is defective for siderophore-mediated iron acquisition. Indeed, the hemB SCV demonstrated limited utilization of endogenous staphyloferrin B or exogenously provided staphyloferrin A, deferoxamine mesylate (Desferal), and epinephrine. Direct measurement of intracellular ATP in hemB and WT S. aureus revealed that both strains can generate comparable levels of ATP during exponential growth, suggesting defects in ATP production cannot account for the inability to efficiently utilize siderophores. Defective siderophore utilization by hemB bacteria was also evident in vivo, as administration of Desferal failed to promote hemB bacterial growth in every organ analyzed except for the kidneys. In support of the hypothesis that S. aureus accesses heme in kidney abscesses, in vitro analyses revealed that increased hemin availability enables hemB bacteria to utilize siderophores for growth when iron availability is restricted. Taken together, our data support the conclusion that hemin is used not only as an iron source itself but also as a nutrient that promotes utilization of siderophore-iron complexes. IMPORTANCE S. aureus small-colony variants (SCVs) are associated with chronic recurrent infection and worsened clinical outcome. SCVs persist within the host despite administration of antibiotics. This study yields insight into how S. aureus SCVs acquire iron, which during infection of a host is a difficult-to-acquire metal nutrient. Under hemin-limited conditions, hemB S. aureus is impaired for siderophore-dependent growth, and in agreement, murine infection indicates that hemin-deficient SCVs meet their nutritional requirement for iron through utilization of hemin. Importantly, we demonstrate that hemB SCVs rely upon hemin as a nutrient to promote siderophore utilization. Therefore, perturbation of heme biosynthesis and/or utilization represents a viable to strategy to mitigate the ability of SCV bacteria to acquire siderophore-bound iron during infection.
Keywords: SCV; heme; iron starvation; pathogenesis; siderophores.
Figures

References
-
- Kochanek KD, Xu J, Murphy SL, Miniño AM, Kung H-C. 2012. Deaths: final data for 2009. Natl Vital Stat Rep 60:1–116. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
