

Canary in a coal mine: collided ribosomes as sensors of cellular conditions
- PMID: 34607755
- PMCID: PMC8688274
- DOI: 10.1016/j.tibs.2021.09.001
Canary in a coal mine: collided ribosomes as sensors of cellular conditions
Abstract
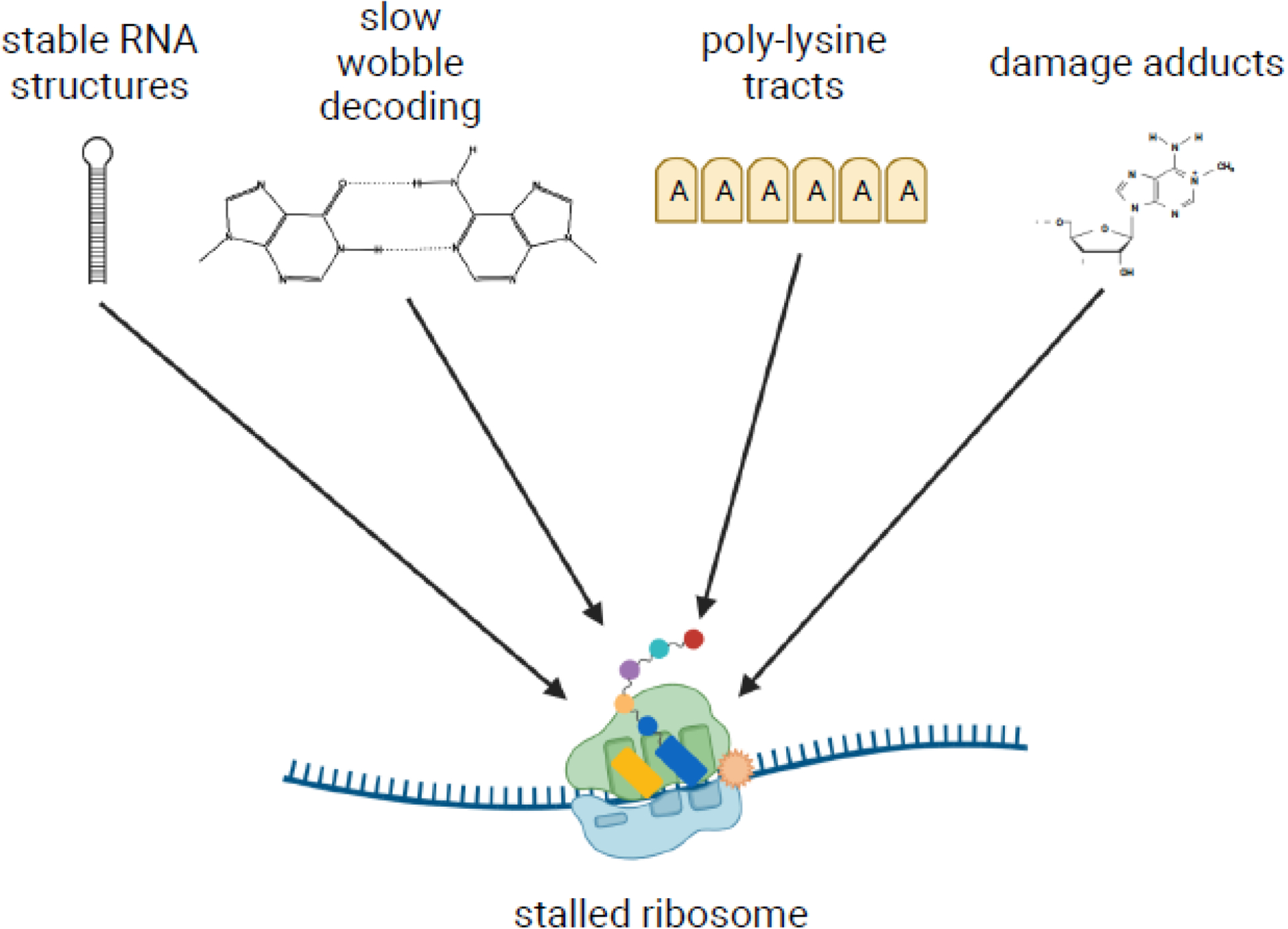
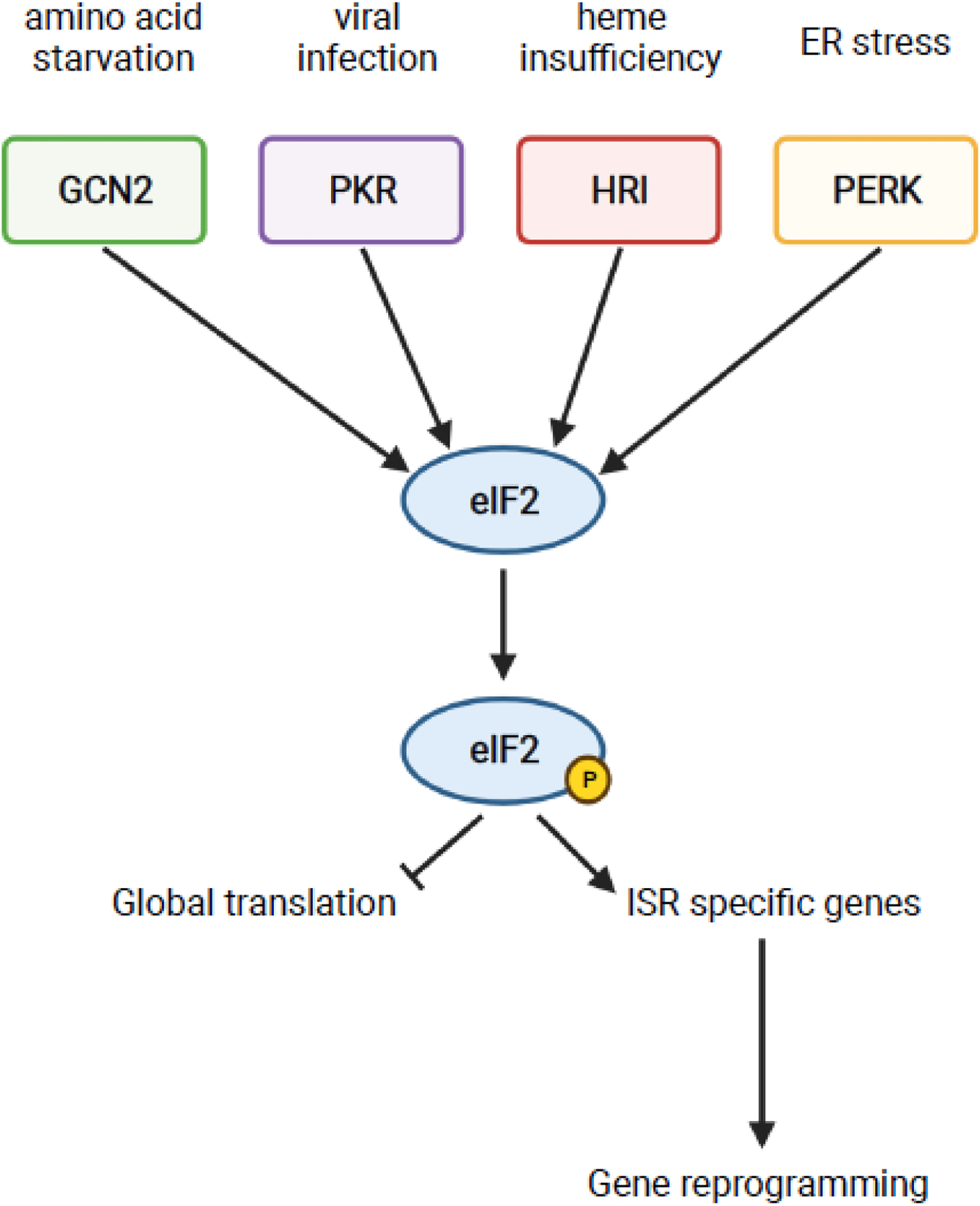
The recent discovery that collision of ribosomes triggers quality control and stress responses in eukaryotes has shifted the perspective of the field. Collided eukaryotic ribosomes adopt a unique structure, acting as a ubiquitin signaling platform for various response factors. While several of the signals that determine which downstream pathways are activated have been uncovered, we are only beginning to learn how the specificity for the activation of each process is achieved during collisions. This review will summarize those findings and how ribosome-associated factors act as molecular sentinels, linking aberrations in translation to the overall cellular state. Insights into how cells respond to ribosome collision events will provide greater understanding of the role of the ribosome in the maintenance of cellular homeostasis.
Keywords: collisions; integrated stress response; mRNA surveillance; quality control; ribosome; signaling.
Copyright © 2021 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Declaration of interests None are declared.
Figures
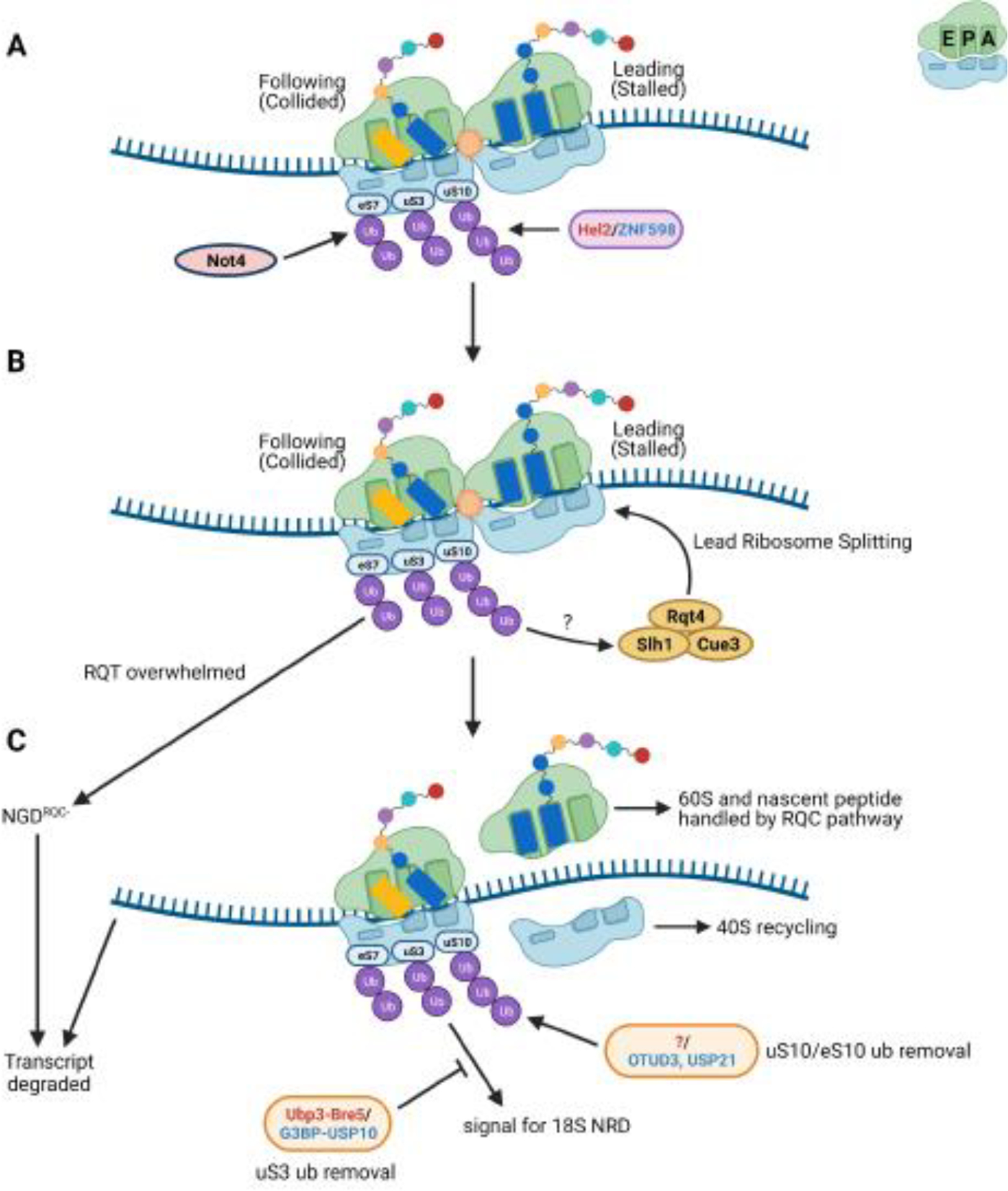
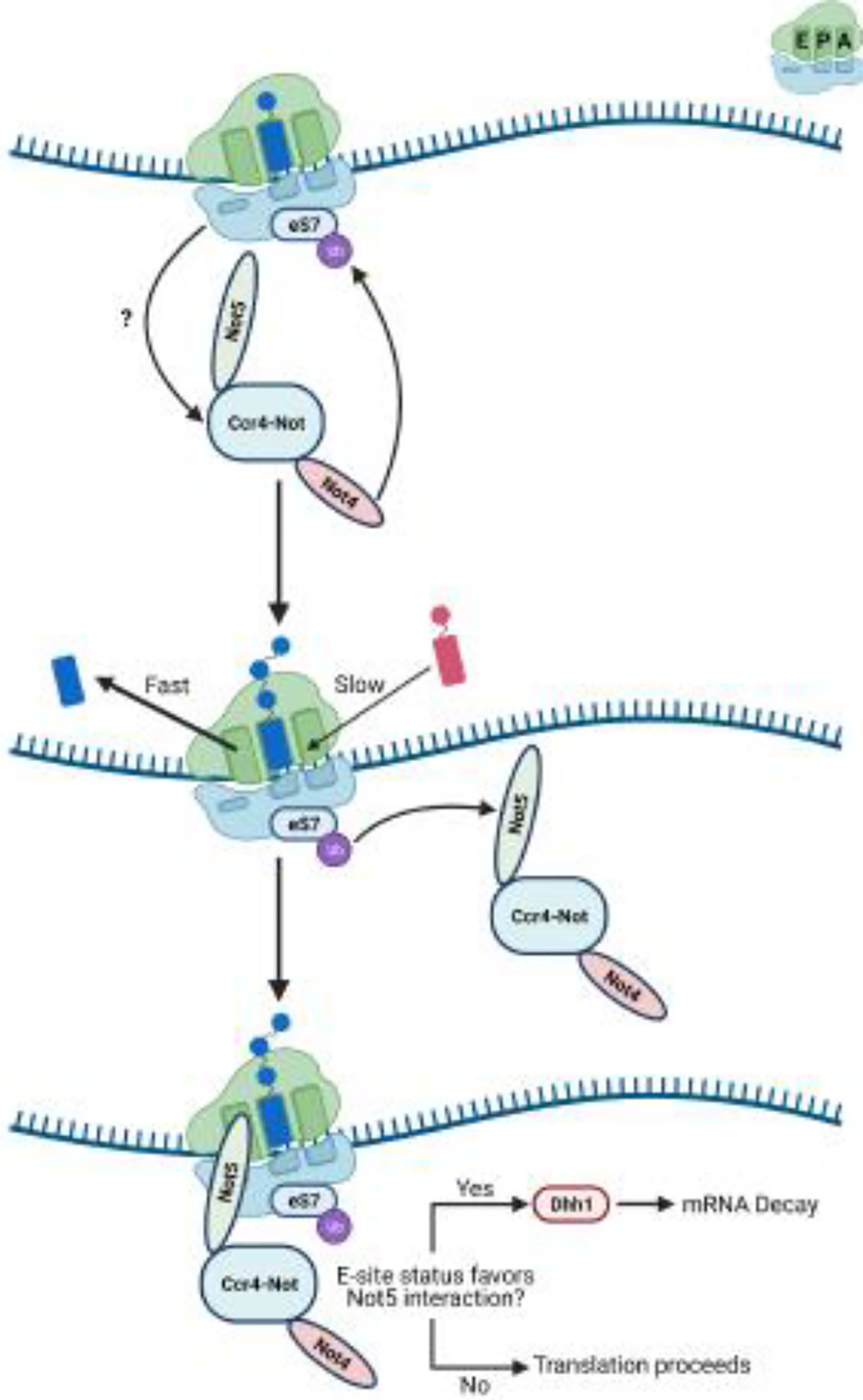
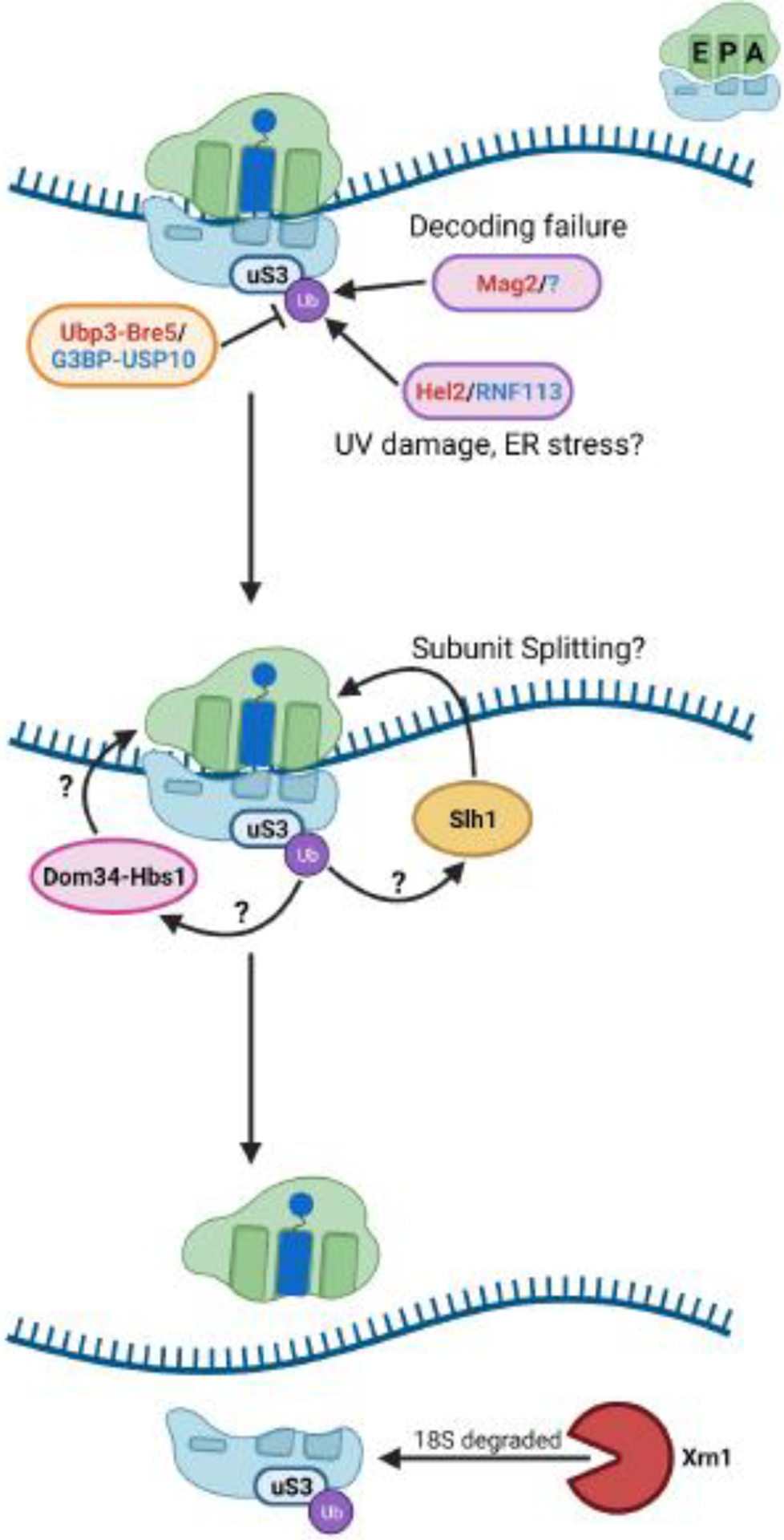
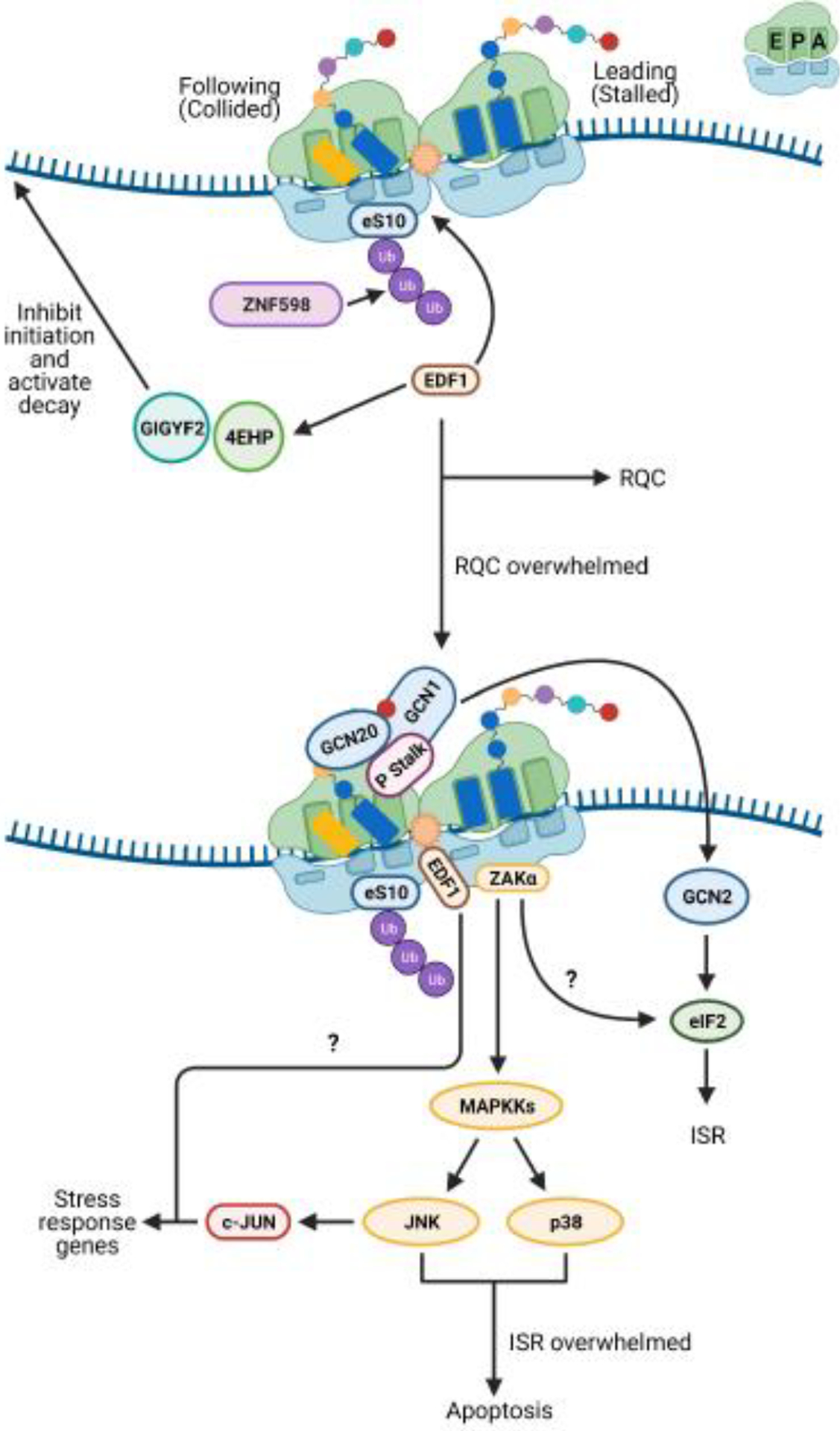
References
-
- García-Muse T and Aguilera A (2016) Transcription-replication conflicts: How they occur and how they are resolved. Nat. Rev. Mol. Cell Biol 17, 553–563 - PubMed
-
- Sørensen MA and Pedersen S (1991) Absolute in vivo translation rates of individual codons in Escherichia coli. The two glutamic acid codons GAA and GAG are translated with a threefold difference in rate. J. Mol. Biol 222, 265–280 - PubMed
-
- Mitarai N et al. (2008) Ribosome Collisions and Translation Efficiency: Optimization by Codon Usage and mRNA Destabilization. J. Mol. Biol 382, 236–245 - PubMed
-
- Farabaugh PJ (1996) Programmed translational frameshifting. Annu. Rev. Genet 30, 507–528 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
