

Metabolic and epigenetic regulation of endoderm differentiation
- PMID: 34607773
- PMCID: PMC8760149
- DOI: 10.1016/j.tcb.2021.09.002
Metabolic and epigenetic regulation of endoderm differentiation
Abstract
The endoderm, one of the three primary germ layers, gives rise to lung, liver, stomach, intestine, colon, pancreas, bladder, and thyroid. These endoderm-originated organs are subject to many life-threatening diseases. However, primary cells/tissues from endodermal organs are often difficult to grow in vitro. Human pluripotent stem cells (hPSCs), therefore, hold great promise for generating endodermal cells and their derivatives for the development of new therapeutics against these human diseases. Although a wealth of research has provided crucial information on the mechanisms underlying endoderm differentiation from hPSCs, increasing evidence has shown that metabolism, in connection with epigenetics, actively regulates endoderm differentiation in addition to the conventional endoderm inducing signals. Here we review recent advances in metabolic and epigenetic regulation of endoderm differentiation.
Keywords: endoderm differentiation; endodermal gene expression; epigenetic remodeling; histone crotonylation; metabolic switch.
Published by Elsevier Ltd.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
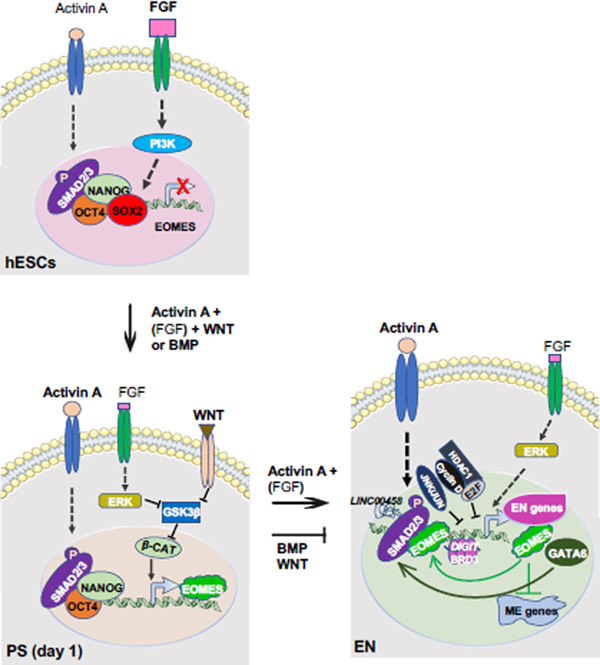
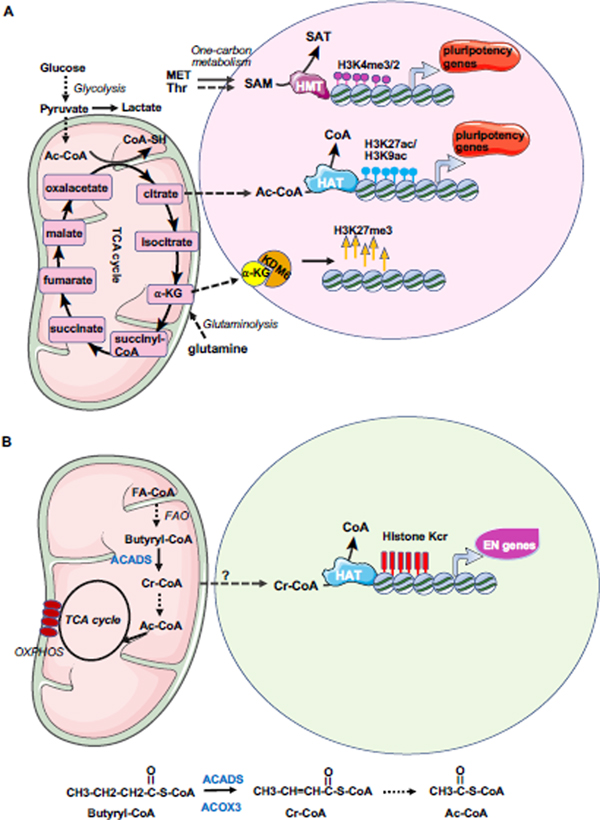
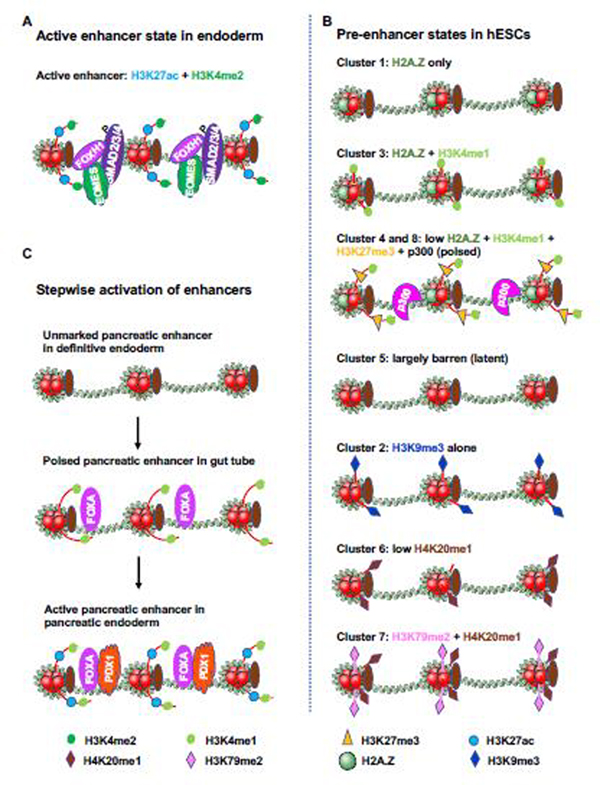
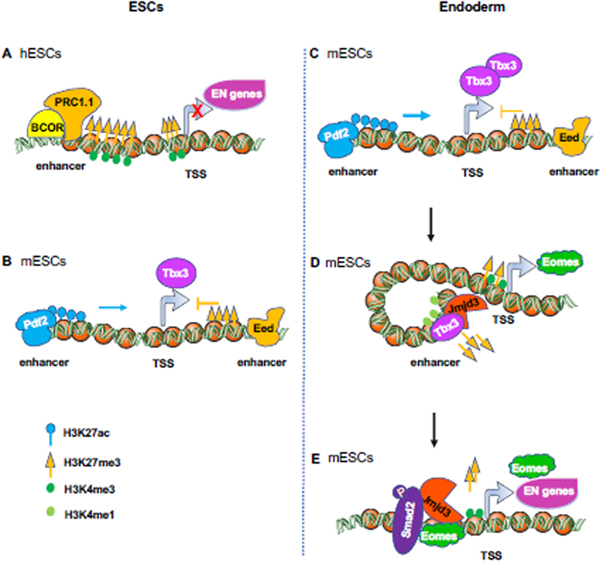
References
-
- Yiangou L et al. (2018) Human Pluripotent Stem Cell-Derived Endoderm for Modeling Development and Clinical Applications. Cell Stem Cell 22 (4), 485–499. - PubMed
-
- Tam PP and Beddington RS (1987) The formation of mesodermal tissues in the mouse embryo during gastrulation and early organogenesis. Development 99 (1), 109–26. - PubMed
-
- Lawson KA et al. (1991) Clonal analysis of epiblast fate during germ layer formation in the mouse embryo. Development 113 (3), 891–911. - PubMed
-
- Tada S et al. (2005) Characterization of mesendoderm: a diverging point of the definitive endoderm and mesoderm in embryonic stem cell differentiation culture. Development 132 (19), 4363–74. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
