

The interplay of immunology and cachexia in infection and cancer
- PMID: 34608281
- PMCID: PMC8489366
- DOI: 10.1038/s41577-021-00624-w
The interplay of immunology and cachexia in infection and cancer
Abstract
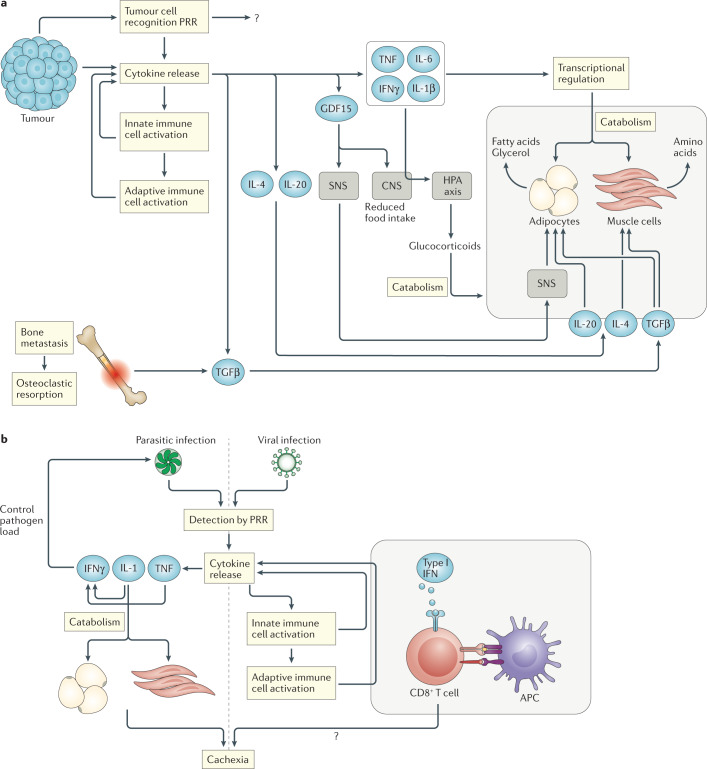
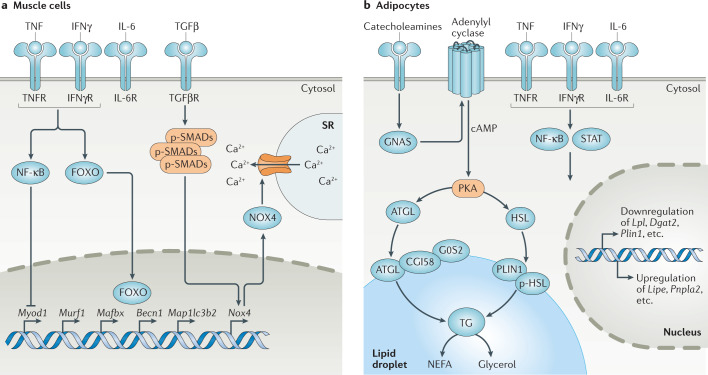
Diverse inflammatory diseases, infections and malignancies are associated with wasting syndromes. In many of these conditions, the standards for diagnosis and treatment are lacking due to our limited understanding of the causative molecular mechanisms. Here, we discuss the complex immunological context of cachexia, a systemic catabolic syndrome that depletes both fat and muscle mass with profound consequences for patient prognosis. We highlight the main cytokine and immune cell-driven pathways that have been linked to weight loss and tissue wasting in the context of cancer-associated and infection-associated cachexia. Moreover, we discuss the potential immunometabolic consequences of cachexia on the basis of newly identified pathways and explore the multilayered area of immunometabolic crosstalk both upstream and downstream of tissue catabolism. Collectively, this Review highlights the intricate relationship of the immune system with cachexia in the context of malignant and infectious diseases.
© 2021. Springer Nature Limited.
Conflict of interest statement
The authors declare no competing interests.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
