

A STING inhibitor suppresses EBV-induced B cell transformation and lymphomagenesis
- PMID: 34609775
- PMCID: PMC8645724
- DOI: 10.1111/cas.15152
A STING inhibitor suppresses EBV-induced B cell transformation and lymphomagenesis
Abstract
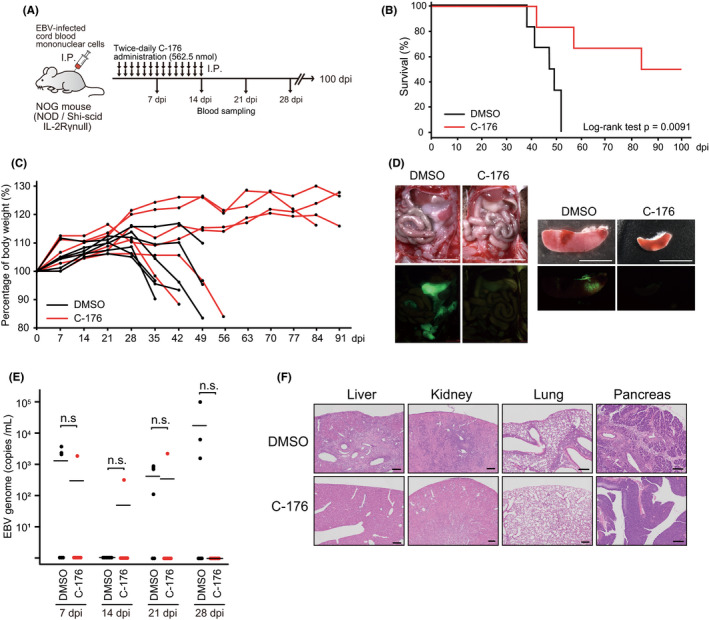
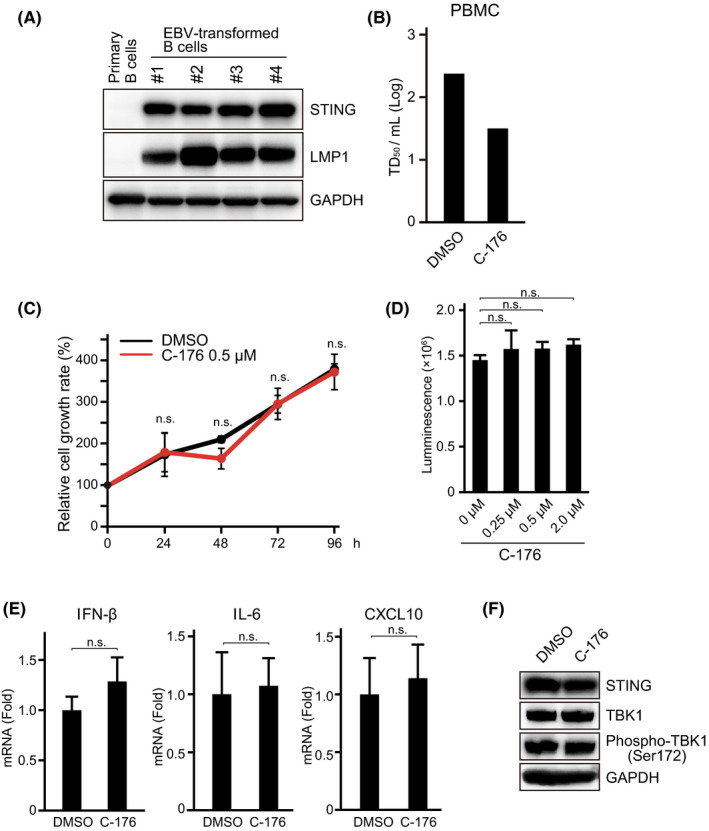
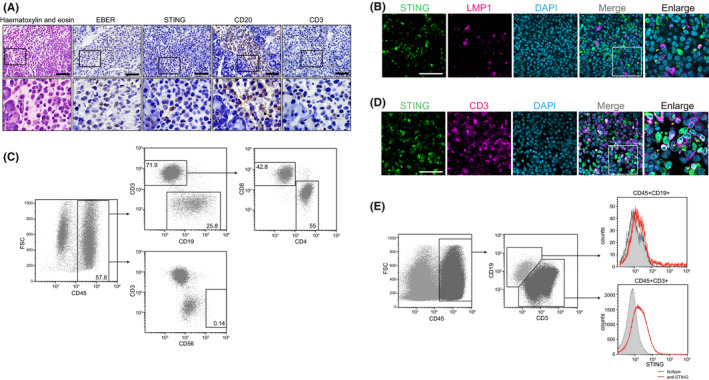
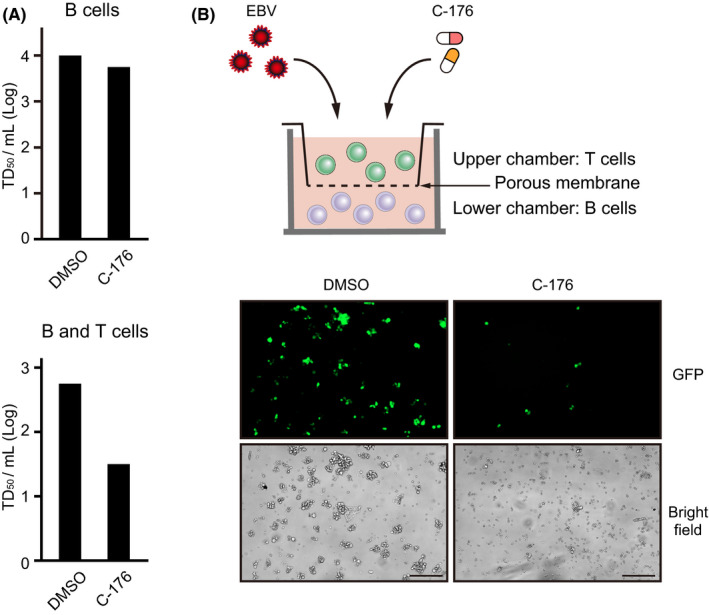
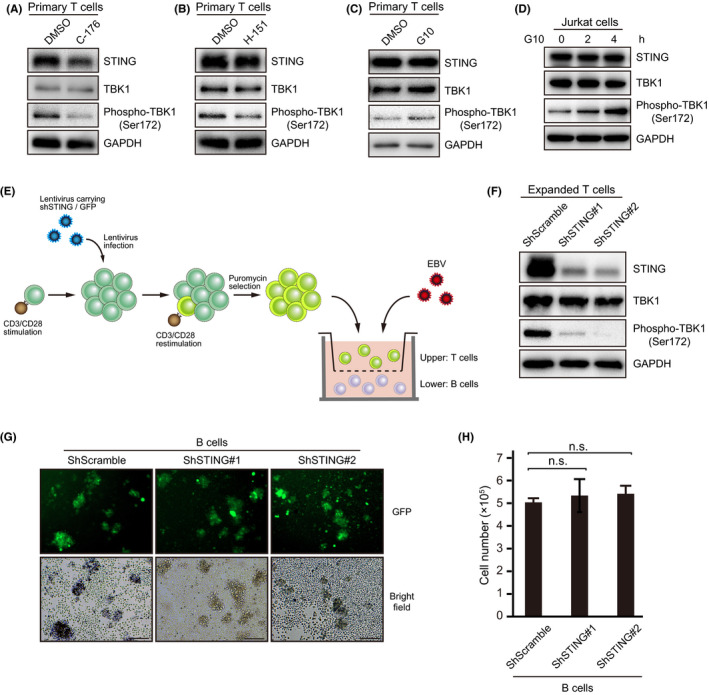
Epstein-Barr virus-associated lymphoproliferative disease (EBV-LPD) is frequently fatal. Innate immunity plays a key role in protecting against pathogens and cancers. The stimulator of interferon genes (STING) is regarded as a key adaptor protein allowing DNA sensors recognizing exogenous cytosolic DNA to activate the type I interferon signaling cascade. In terms of EBV tumorigenicity, the role of STING remains elusive. Here we showed that treatment with the STING inhibitor, C-176, suppressed EBV-induced transformation in peripheral blood mononuclear cells. In an EBV-LPD mouse model, C-176 treatment also inhibited tumor formation and prolonged survival. Treatment with B cells alone did not affect EBV transformation, but suppression of EBV-induced transformation was observed in the presence of T cells. Even without direct B cell-T cell contact in a transwell system, the inhibitor reduced the transformation activity, indicating that intercellular communication by humoral factors was critical to prevent EBV-induced transformation. These findings suggest that inhibition of STING signaling pathway with C-176 could be a new therapeutic target of EBV-LPD.
Keywords: EBV-LPD; Epstein-Barr virus; PAMPs; STING; cGAS.
© 2021 The Authors. Cancer Science published by John Wiley & Sons Australia, Ltd on behalf of Japanese Cancer Association.
Conflict of interest statement
H. Kimura and Y. Sato were supported by grants from Bristol Myers Squibb and Merck Sharp & Dohme, respectively. All other authors have no conflicts of interest directly relevant to the content of this article.
Figures
Similar articles
-
The translation inhibitor silvestrol exhibits direct anti-tumor activity while preserving innate and adaptive immunity against EBV-driven lymphoproliferative disease.Oncotarget. 2015 Feb 20;6(5):2693-708. doi: 10.18632/oncotarget.2098. Oncotarget. 2015. PMID: 25393910 Free PMC article.
-
Epstein-Barr virus (EBV) and lymphomagenesis.Front Biosci. 2002 Feb 1;7:e58-65. doi: 10.2741/A907. Front Biosci. 2002. PMID: 11815280 Review.
-
Leflunomide/teriflunomide inhibit Epstein-Barr virus (EBV)- induced lymphoproliferative disease and lytic viral replication.Oncotarget. 2017 Jul 4;8(27):44266-44280. doi: 10.18632/oncotarget.17863. Oncotarget. 2017. PMID: 28574826 Free PMC article.
-
Anti-tumor effects of suberoylanilide hydroxamic acid on Epstein-Barr virus-associated T cell and natural killer cell lymphoma.Cancer Sci. 2014 Jun;105(6):713-22. doi: 10.1111/cas.12418. Epub 2014 May 13. Cancer Sci. 2014. PMID: 24712440 Free PMC article.
-
B lymphocytes and Epstein-Barr virus: the lesson of post-transplant lymphoproliferative disorders.Autoimmun Rev. 2007 Dec;7(2):96-101. doi: 10.1016/j.autrev.2007.02.012. Epub 2007 Mar 26. Autoimmun Rev. 2007. PMID: 18035317 Review.
Cited by
-
Epstein-Barr Virus BBLF1 Mediates Secretory Vesicle Transport to Facilitate Mature Virion Release.J Virol. 2023 Jun 29;97(6):e0043723. doi: 10.1128/jvi.00437-23. Epub 2023 May 17. J Virol. 2023. PMID: 37195206 Free PMC article.
-
The cyclic guanosine monophosphate synthase-stimulator of interferon genes pathway as a potential target for tumor immunotherapy.Front Immunol. 2023 Apr 21;14:1121603. doi: 10.3389/fimmu.2023.1121603. eCollection 2023. Front Immunol. 2023. PMID: 37153627 Free PMC article. Review.
-
Multi-omics integration reveals the oncogenic role of eccDNAs in diffuse large B-cell lymphoma through STING signalling.Clin Transl Med. 2024 Aug;14(8):e1815. doi: 10.1002/ctm2.1815. Clin Transl Med. 2024. PMID: 39183480 Free PMC article.
-
Growth Transformation of B Cells by Epstein-Barr Virus Requires IMPDH2 Induction and Nucleolar Hypertrophy.Microbiol Spectr. 2023 Aug 17;11(4):e0044023. doi: 10.1128/spectrum.00440-23. Epub 2023 Jul 6. Microbiol Spectr. 2023. PMID: 37409959 Free PMC article.
-
Prevalence of EBV, HHV6, HCMV, HAdV, SARS-CoV-2, and Autoantibodies to Type I Interferon in Sputum from Myalgic Encephalomyelitis/Chronic Fatigue Syndrome Patients.Viruses. 2025 Mar 14;17(3):422. doi: 10.3390/v17030422. Viruses. 2025. PMID: 40143349 Free PMC article.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
