

Aging impairs human bone marrow function and cardiac repair following myocardial infarction in a humanized chimeric mouse
- PMID: 34612564
- PMCID: PMC8590094
- DOI: 10.1111/acel.13494
Aging impairs human bone marrow function and cardiac repair following myocardial infarction in a humanized chimeric mouse
Abstract
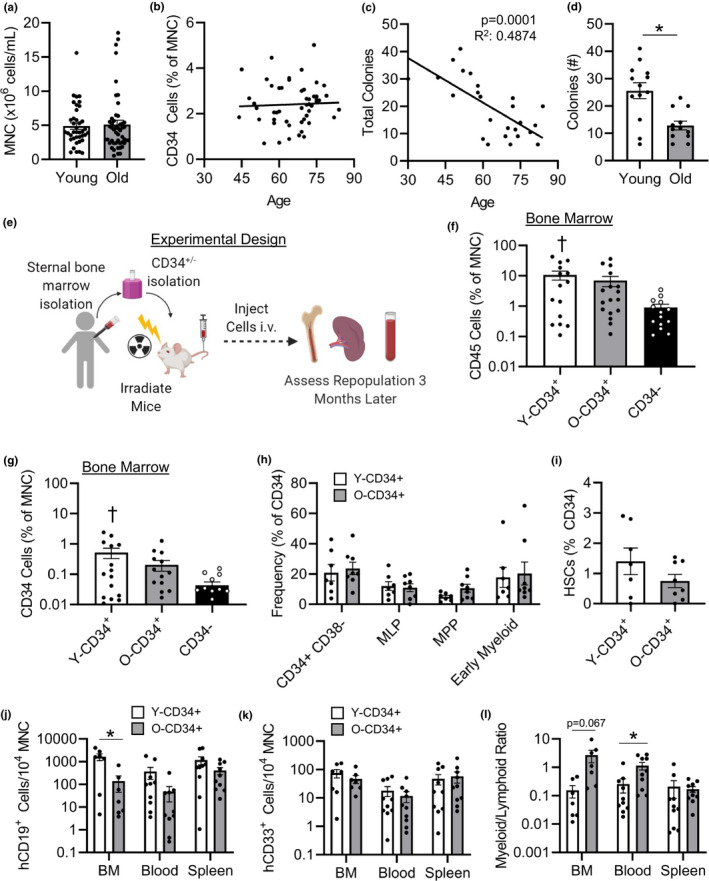
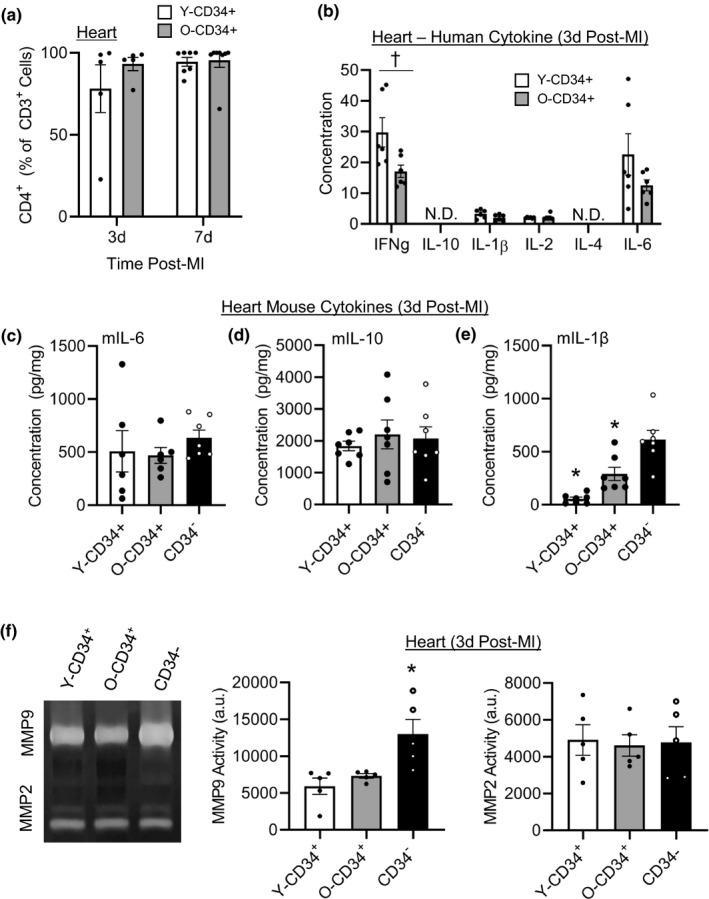
Ventricular remodeling following myocardial infarction (MI) is a major cause of heart failure, a condition prevalent in older individuals. Following MI, immune cells are mobilized to the myocardium from peripheral lymphoid organs and play an active role in orchestrating repair. While the effect of aging on mouse bone marrow (BM) has been studied, less is known about how aging affects human BM cells and their ability to regulate repair processes. In this study, we investigate the effect aging has on human BM cell responses post-MI using a humanized chimeric mouse model. BM samples were collected from middle aged (mean age 56.4 ± 0.97) and old (mean age 72.7 ± 0.59) patients undergoing cardiac surgery, CD34+/- cells were isolated, and NOD-scid-IL2rγnull (NSG) mice were reconstituted. Three months following reconstitution, the animals were examined at baseline or subjected to coronary artery ligation (MI). Younger patient cells exhibited greater repopulation capacity in the BM, blood, and spleen as well as greater lymphoid cell production. Following MI, CD34+ cell age impacted donor and host cellular responses. Mice reconstituted with younger CD34+ cells exhibited greater human CD45+ recruitment to the heart compared to mice reconstituted with old cells. Increased cellular responses were primarily driven by T-cell recruitment, and these changes corresponded with greater human IFNy levels and reduced mouse IL-1β in the heart. Age-dependent changes in BM function led to significantly lower survival, increased infarct expansion, impaired host cell responses, and reduced function by 4w post-MI. In contrast, younger CD34+ cells helped to limit remodeling and preserve function post-MI.
Keywords: aging; bone marrow transplant; humanized mice; myocardial infarction.
© 2021 The Authors. Aging Cell published by Anatomical Society and John Wiley & Sons Ltd.
Conflict of interest statement
None.
Figures




References
-
- Alibhai, F. J. , Lim, F. , Yeganeh, A. , DiStefano, P. V. , Binesh‐Marvasti, T. , Belfiore, A. , Wlodarek, L. , Gustafson, D. , Millar, S. , Li, S.‐H. , Weisel, R. D. , Fish, J. E. , & Li, R.‐K. (2020). Cellular senescence contributes to age‐dependent changes in circulating extracellular vesicle cargo and function. Aging Cell, 19(3), e13103. 10.1111/acel.13103 - DOI - PMC - PubMed
-
- Broxmeyer, H. E. , Liu, Y. , Kapur, R. , Orschell, C. M. , Aljoufi, A. , Ropa, J. P. , Trinh, T. , Burns, S. , & Capitano, M. L. (2020). Fate of hematopoiesis during aging. What do we really know, and what are its implications? Stem Cell Reviews and Reports, 16(6), 1020–1048. 10.1007/s12015-020-10065-y - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
