

Aromatic Dimer Dehydrogenases from Novosphingobium aromaticivorans Reduce Monoaromatic Diketones
- PMID: 34613756
- PMCID: PMC8612281
- DOI: 10.1128/AEM.01742-21
Aromatic Dimer Dehydrogenases from Novosphingobium aromaticivorans Reduce Monoaromatic Diketones
Abstract
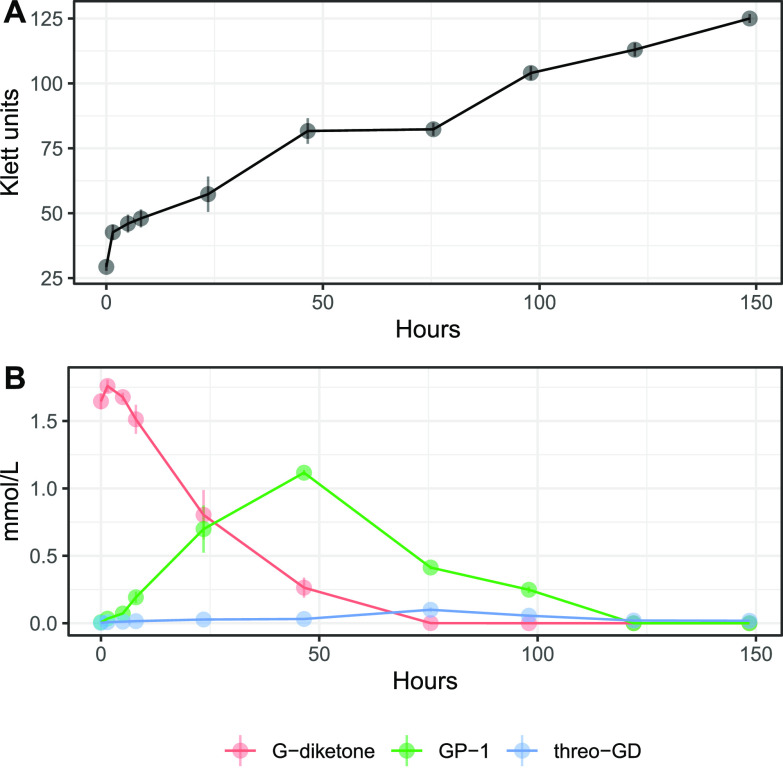
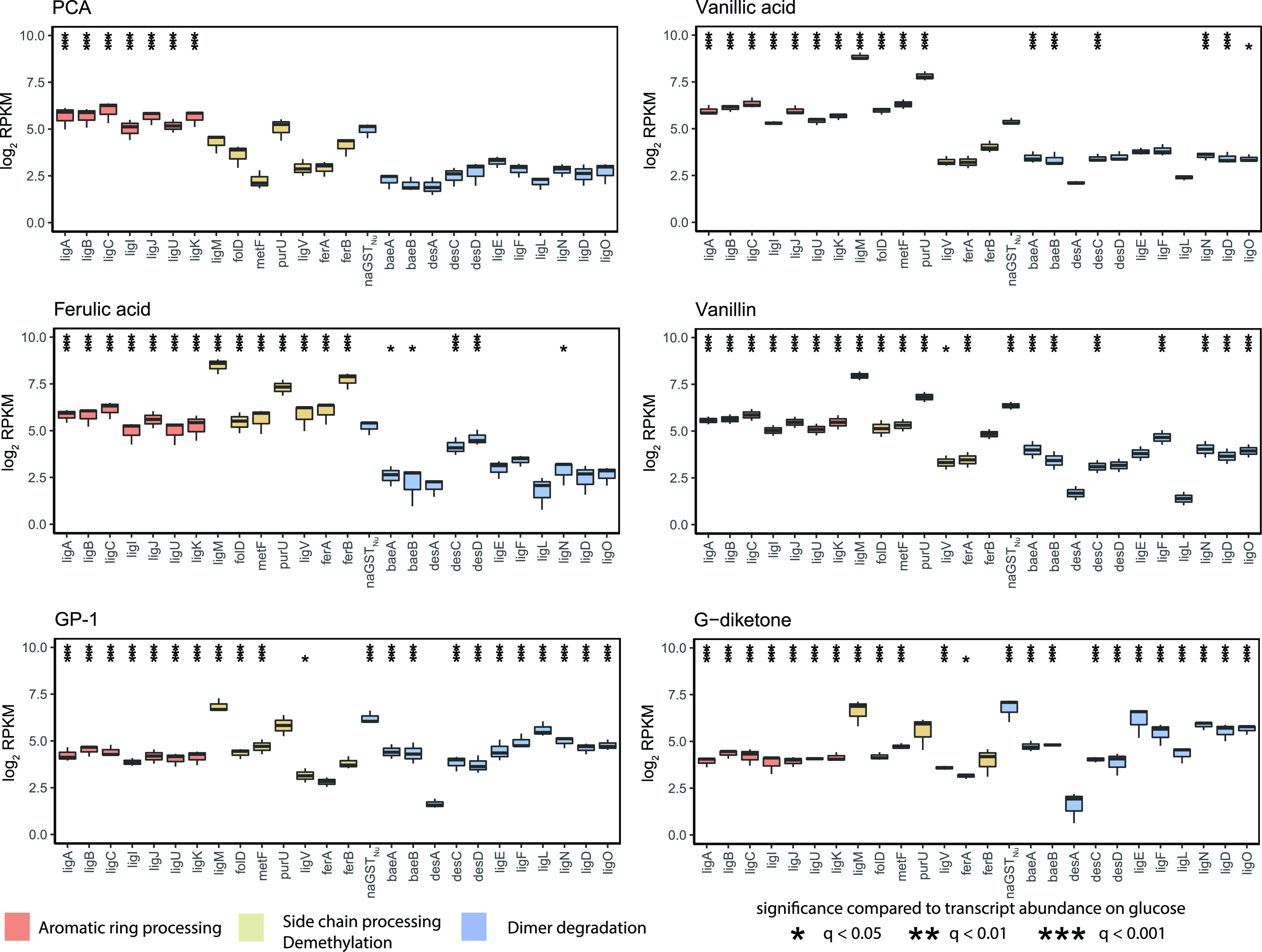
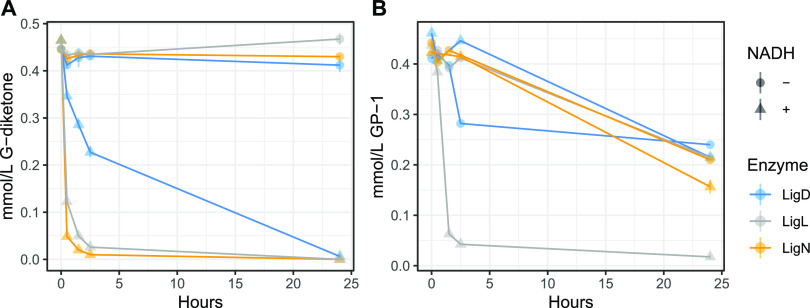
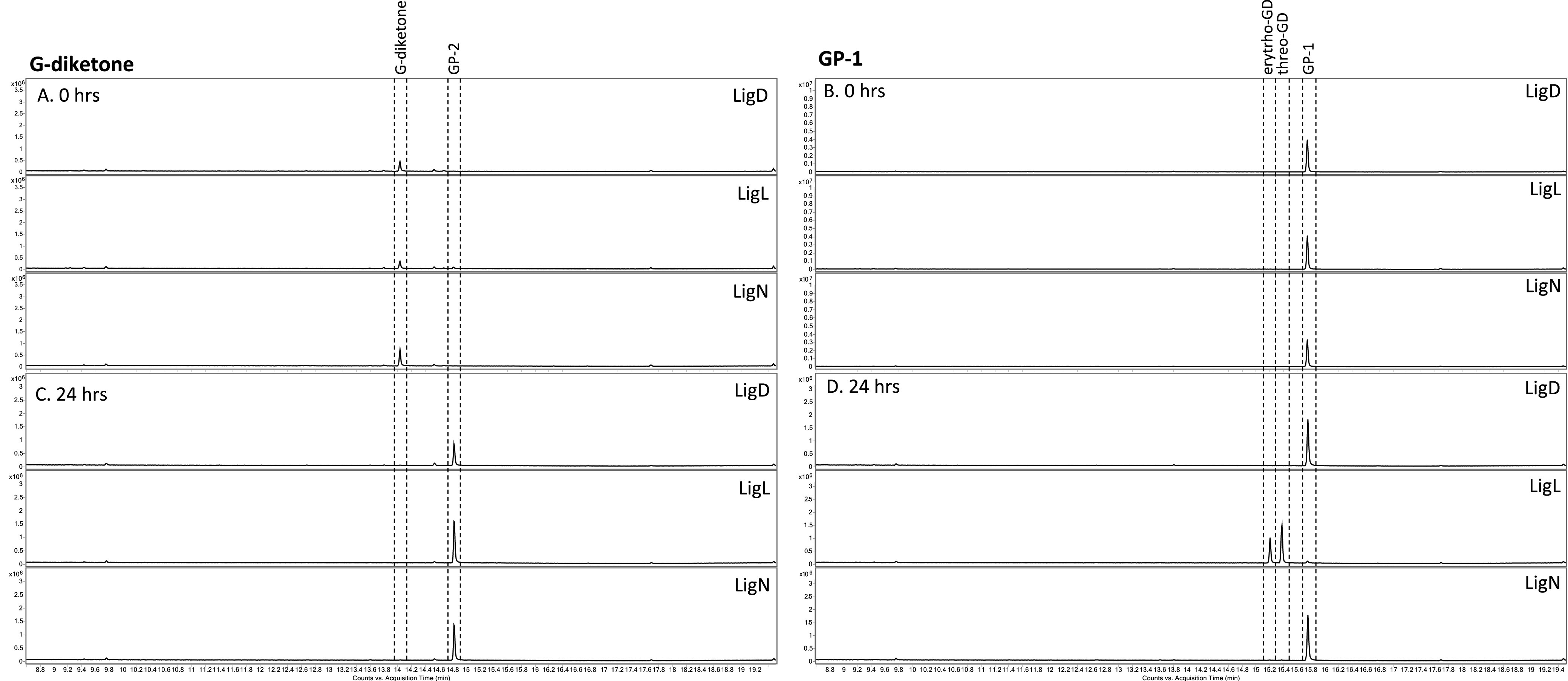
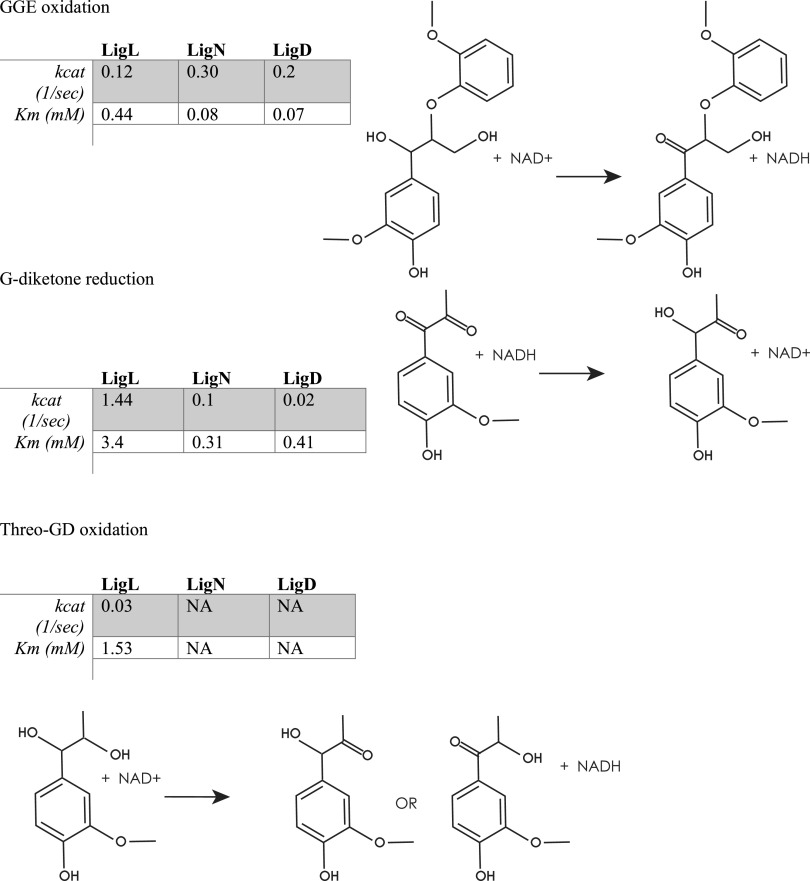
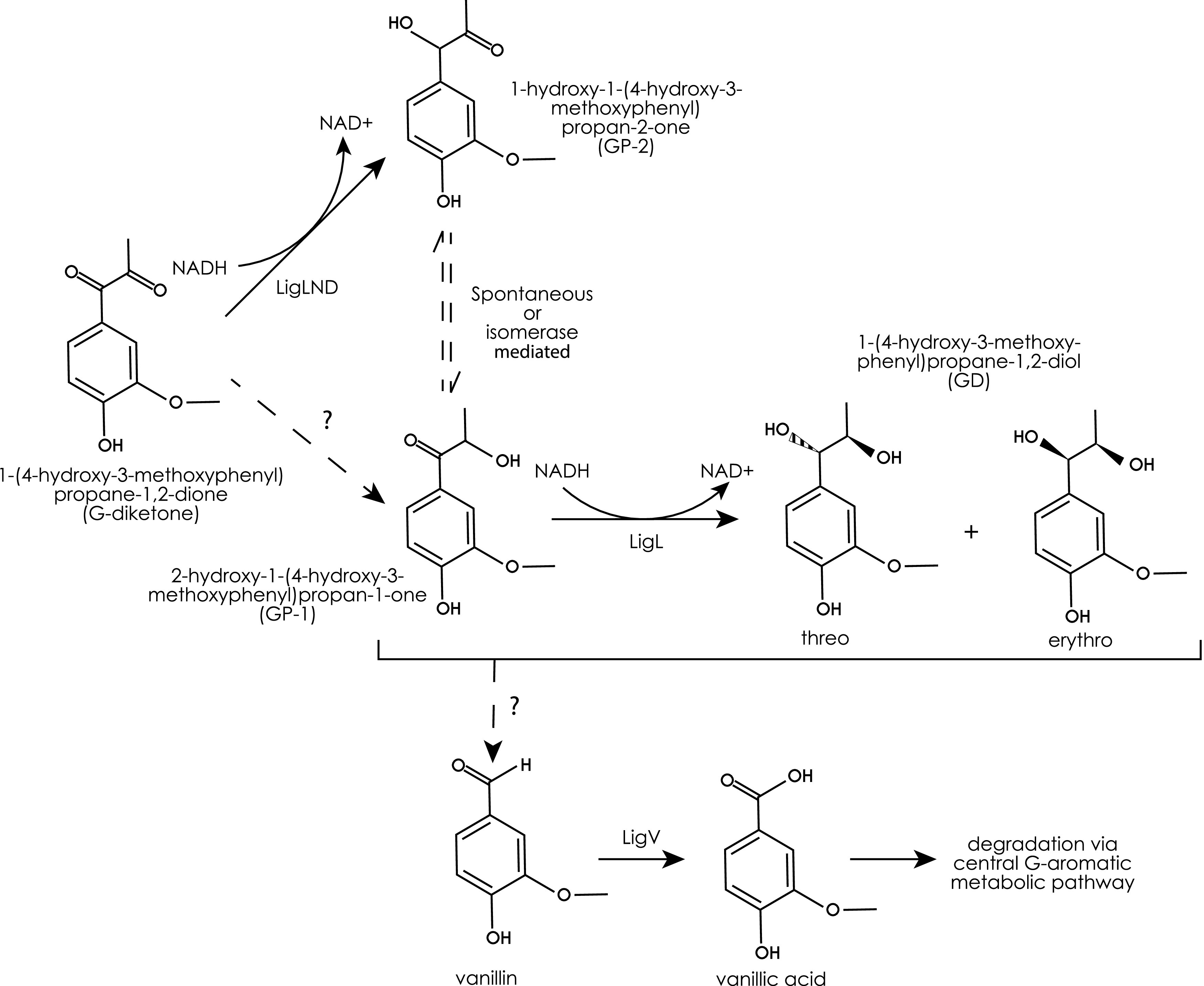
Lignin is a potential source of valuable chemicals, but its chemical depolymerization results in a heterogeneous mixture of aromatics and other products. Microbes could valorize depolymerized lignin by converting multiple substrates into one or a small number of products. In this study, we describe the ability of Novosphingobium aromaticivorans to metabolize 1-(4-hydroxy-3-methoxyphenyl)propane-1,2-dione (G-diketone), an aromatic Hibbert diketone that is produced during formic acid-catalyzed lignin depolymerization. By assaying genome-wide transcript levels from N. aromaticivorans during growth on G-diketone and other chemically-related aromatics, we hypothesized that the Lig dehydrogenases, previously characterized as oxidizing β-O-4 linkages in aromatic dimers, were involved in G-diketone metabolism by N. aromaticivorans. Using purified N. aromaticivorans Lig dehydrogenases, we found that LigL, LigN, and LigD each reduced the Cα ketone of G-diketone in vitro but with different substrate specificities and rates. Furthermore, LigL, but not LigN or LigD, also reduced the Cα ketone of 2-hydroxy-1-(4-hydroxy-3-methoxyphenyl)propan-1-one (GP-1) in vitro, a derivative of G-diketone with the Cβ ketone reduced, when GP-1 was provided as a substrate. The newly identified activity of these Lig dehydrogenases expands the potential range of substrates utilized by N. aromaticivorans beyond what has been previously recognized. This is beneficial both for metabolizing a wide range of natural and non-native depolymerized lignin substrates and for engineering microbes and enzymes that are active with a broader range of aromatic compounds. IMPORTANCE Lignin is a major plant polymer composed of aromatic units that have value as chemicals. However, the structure and composition of lignin have made it difficult to use this polymer as a renewable source of industrial chemicals. Bacteria like Novosphingobium aromaticivorans have the potential to make chemicals from lignin not only because of their natural ability to metabolize a variety of aromatics but also because there are established protocols to engineer N. aromaticivorans strains to funnel lignin-derived aromatics into valuable products. In this work, we report a newly discovered activity of previously characterized dehydrogenase enzymes with a chemically modified by-product of lignin depolymerization. We propose that the activity of N. aromaticivorans enzymes with both native lignin aromatics and those produced by chemical depolymerization will expand opportunities for producing industrial chemicals from the heterogenous components of this abundant plant polymer.
Keywords: Lignin; Novosphingobium; aromatic dehydrogenases; aromatic metabolism; ketone reduction; sphingomonads.
Figures
References
-
- Ragauskas AJ, Beckham GT, Biddy MJ, Chandra R, Chen F, Davis MF, Davison BH, Dixon RA, Gilna P, Keller M, Langan P, Naskar AK, Saddler JN, Tschaplinski TJ, Tuskan GA, Wyman CE. 2014. Lignin valorization: Improving lignin processing in the biorefinery. Science 344:1246843–1246843. 10.1126/science.1246843. - DOI - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
