

Mechanisms of Acetoin Toxicity and Adaptive Responses in an Acetoin-Producing Species, Lactococcus lactis
- PMID: 34613757
- PMCID: PMC8612267
- DOI: 10.1128/AEM.01079-21
Mechanisms of Acetoin Toxicity and Adaptive Responses in an Acetoin-Producing Species, Lactococcus lactis
Abstract
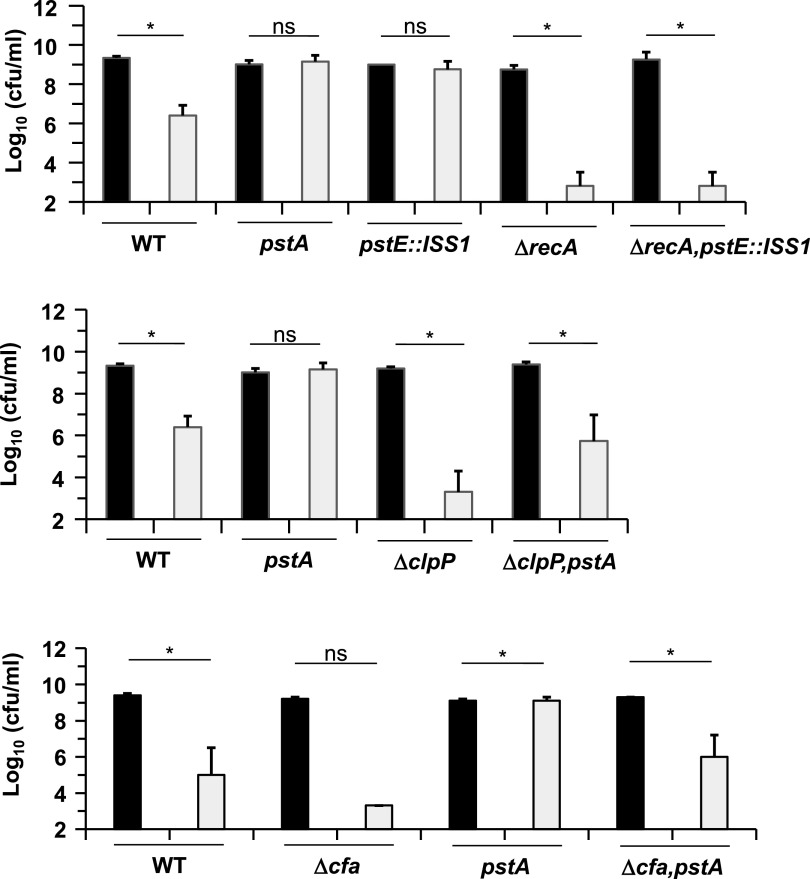
Acetoin, 3-hydroxyl,2-butanone, is extensively used as a flavor additive in food products. This volatile compound is produced by the dairy bacterium Lactococcus lactis when aerobic respiration is activated by haem addition, and comprises ∼70% of carbohydrate degradation products. Here we investigate the targets of acetoin toxicity, and determine how acetoin impacts L. lactis physiology and survival. Acetoin caused damage to DNA and proteins, which related to reactivity of its keto group. Acetoin stress was reflected in proteome profiles, which revealed changes in lipid metabolic proteins. Acetoin provoked marked changes in fatty acid composition, with massive accumulation of cycC19:0 cyclopropane fatty acid at the expense of its unsaturated C18:1 fatty acid precursor. Deletion of the cfa gene, encoding the cycC19:0 synthase, sensitized cells to acetoin stress. Acetoin-resistant transposon mutagenesis revealed a hot spot in the high affinity phosphate transporter operon pstABCDEF, which is known to increase resistance to multiple stresses. This work reveals the causes and consequences of acetoin stress on L. lactis, and may facilitate control of lactic acid bacteria production in technological processes. IMPORTANCE Acetoin, 3-hydroxyl,2-butanone, has diverse uses in chemical industry, agriculture, and dairy industries as a volatile compound that generates aromas. In bacteria, it can be produced in high amount by Lactococcus lactis when it grows under aerobic respiration. However, acetoin production can be toxic and detrimental for growth and/or survival. Our results showed that it damages DNA and proteins via its keto group. We also showed that acetoin modifies membrane fatty acid composition with the production of cyclopropane C19:0 fatty acid at the expense of an unsaturated C18:1. We isolated mutants more resistant to acetoin than the wild-type strain. All of them mapped to a single locus pstABCDEF operon, suggesting a simple means to limit acetoin toxicity in dairy bacteria and to improve its production.
Keywords: Lactococcus lactis; acetoin; fatty acids; pst operon.
Figures







References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
