

Atypical molecular features of RNA silencing against the phloem-restricted polerovirus TuYV
- PMID: 34614168
- PMCID: PMC8565345
- DOI: 10.1093/nar/gkab802
Atypical molecular features of RNA silencing against the phloem-restricted polerovirus TuYV
Abstract
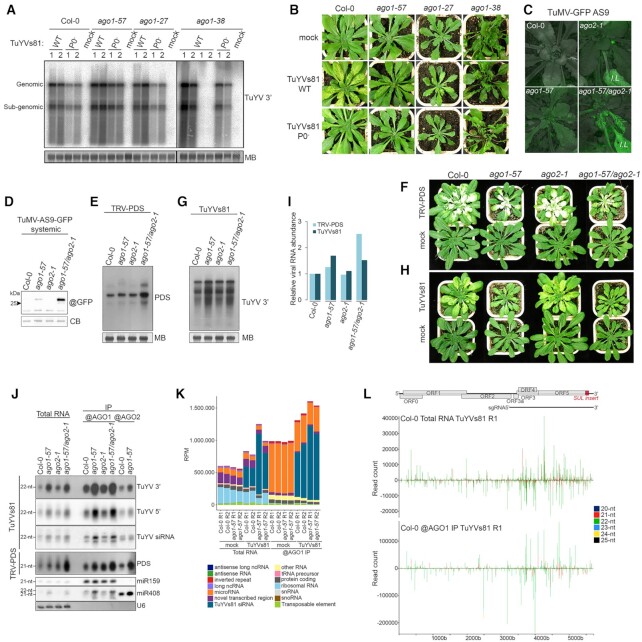
In plants and some animal lineages, RNA silencing is an efficient and adaptable defense mechanism against viruses. To counter it, viruses encode suppressor proteins that interfere with RNA silencing. Phloem-restricted viruses are spreading at an alarming rate and cause substantial reduction of crop yield, but how they interact with their hosts at the molecular level is still insufficiently understood. Here, we investigate the antiviral response against phloem-restricted turnip yellows virus (TuYV) in the model plant Arabidopsis thaliana. Using a combination of genetics, deep sequencing, and mechanical vasculature enrichment, we show that the main axis of silencing active against TuYV involves 22-nt vsiRNA production by DCL2, and their preferential loading into AGO1. Moreover, we identify vascular secondary siRNA produced from plant transcripts and initiated by DCL2-processed AGO1-loaded vsiRNA. Unexpectedly, and despite the viral encoded VSR P0 previously shown to mediate degradation of AGO proteins, vascular AGO1 undergoes specific post-translational stabilization during TuYV infection. Collectively, our work uncovers the complexity of antiviral RNA silencing against phloem-restricted TuYV and prompts a re-assessment of the role of its suppressor of silencing P0 during genuine infection.
© The Author(s) 2021. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures




References
-
- Pumplin N., Voinnet O.. RNA silencing suppression by plant pathogens: defence, counter-defence and counter-counter-defence. Nat. Rev. Microbiol. 2013; 11:745–760. - PubMed
-
- Yang Z., Li Y.. Dissection of RNAi-based antiviral immunity in plants. Curr. Opin. Virol. 2018; 32:88–99. - PubMed
-
- Deleris A., Gallago-Bartolome J., Bao J., Kasschau K.D., Carrington J.C., Voinnet O.. Hierarchical action and inhibition of plant dicer-like proteins in antiviral defense. Science (80-.). 2006; 313:68–71. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
