

Transcription Elongation Machinery Is a Druggable Dependency and Potentiates Immunotherapy in Glioblastoma Stem Cells
- PMID: 34615656
- PMCID: PMC8831451
- DOI: 10.1158/2159-8290.CD-20-1848
Transcription Elongation Machinery Is a Druggable Dependency and Potentiates Immunotherapy in Glioblastoma Stem Cells
Abstract
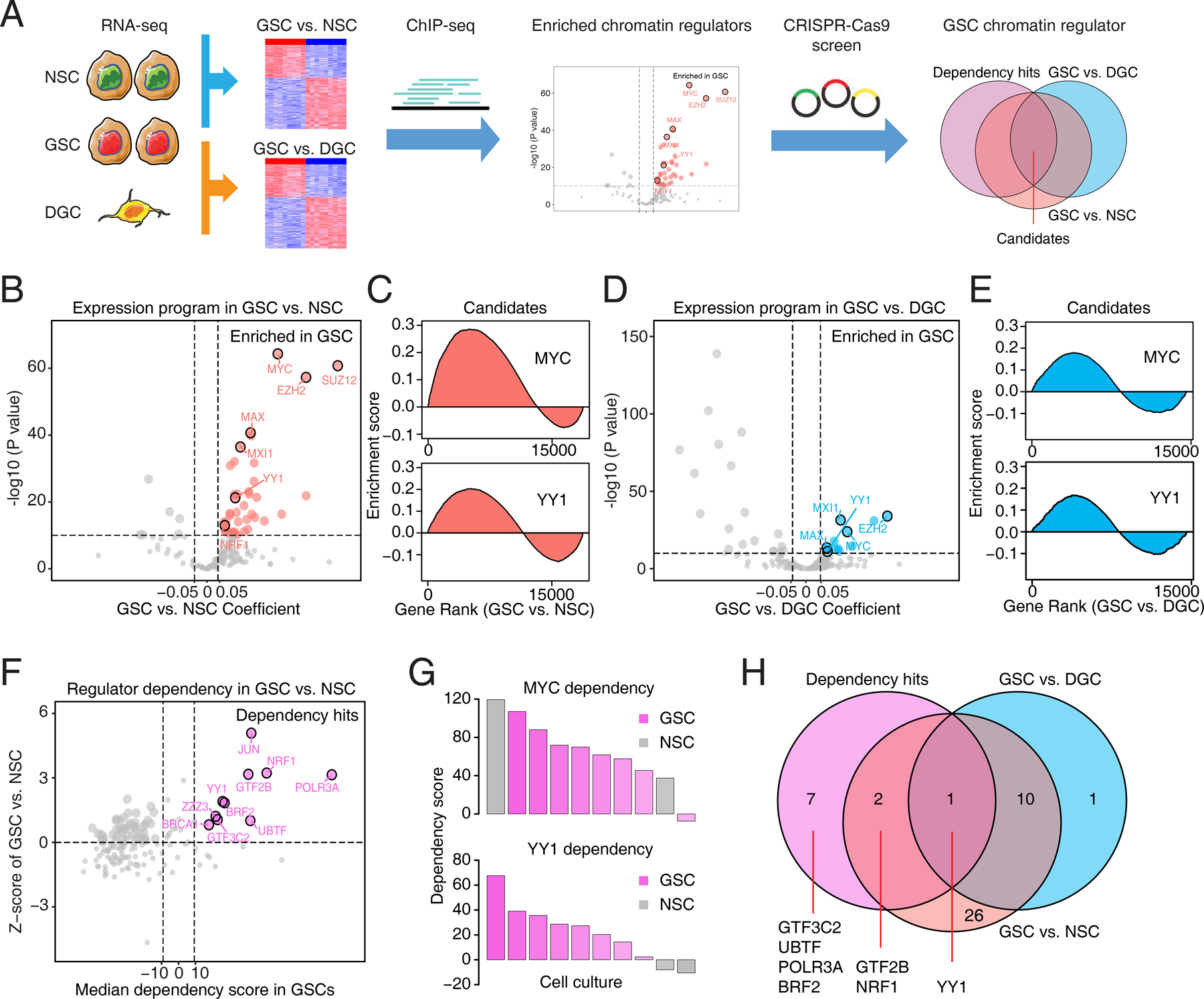
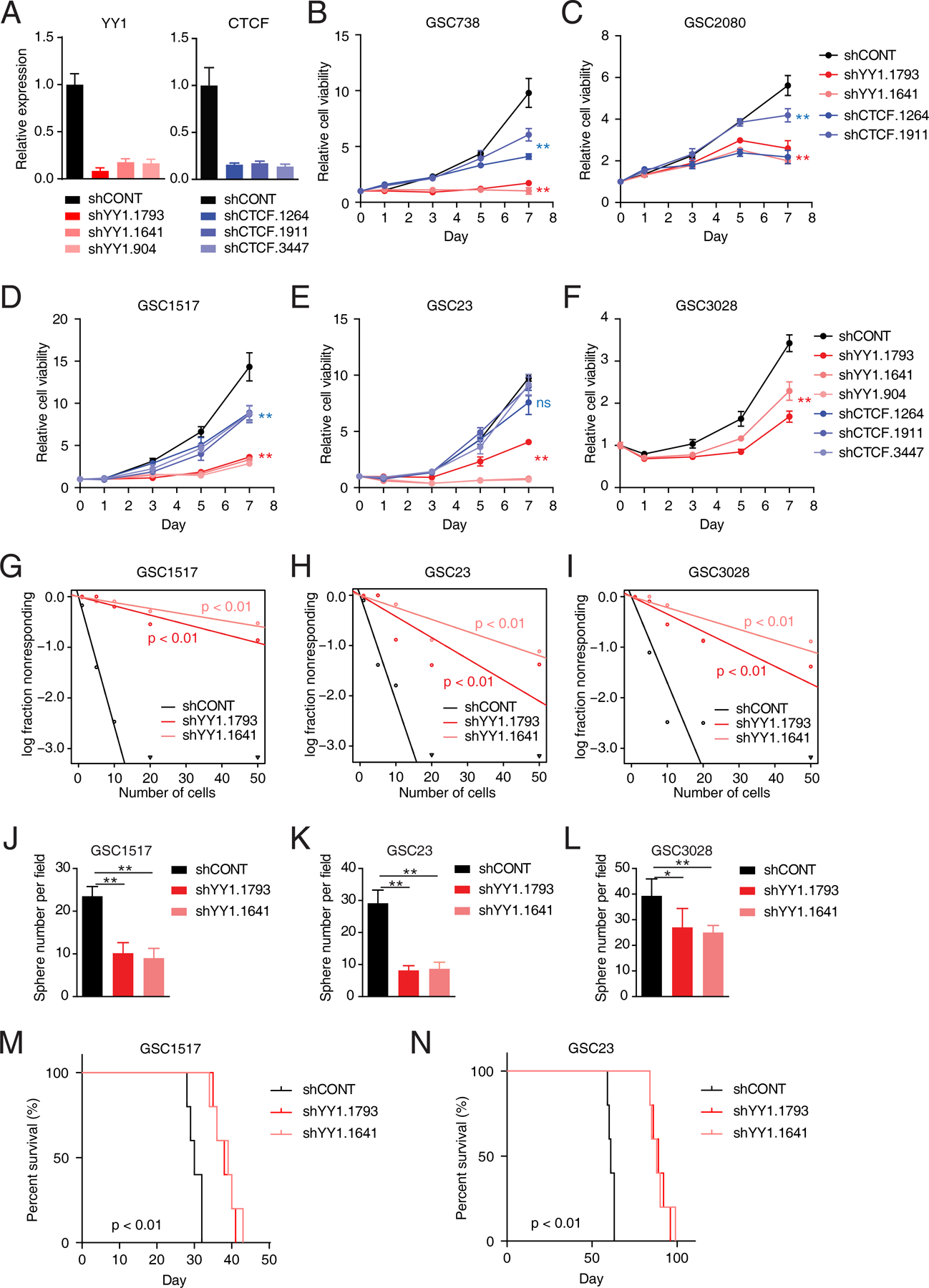
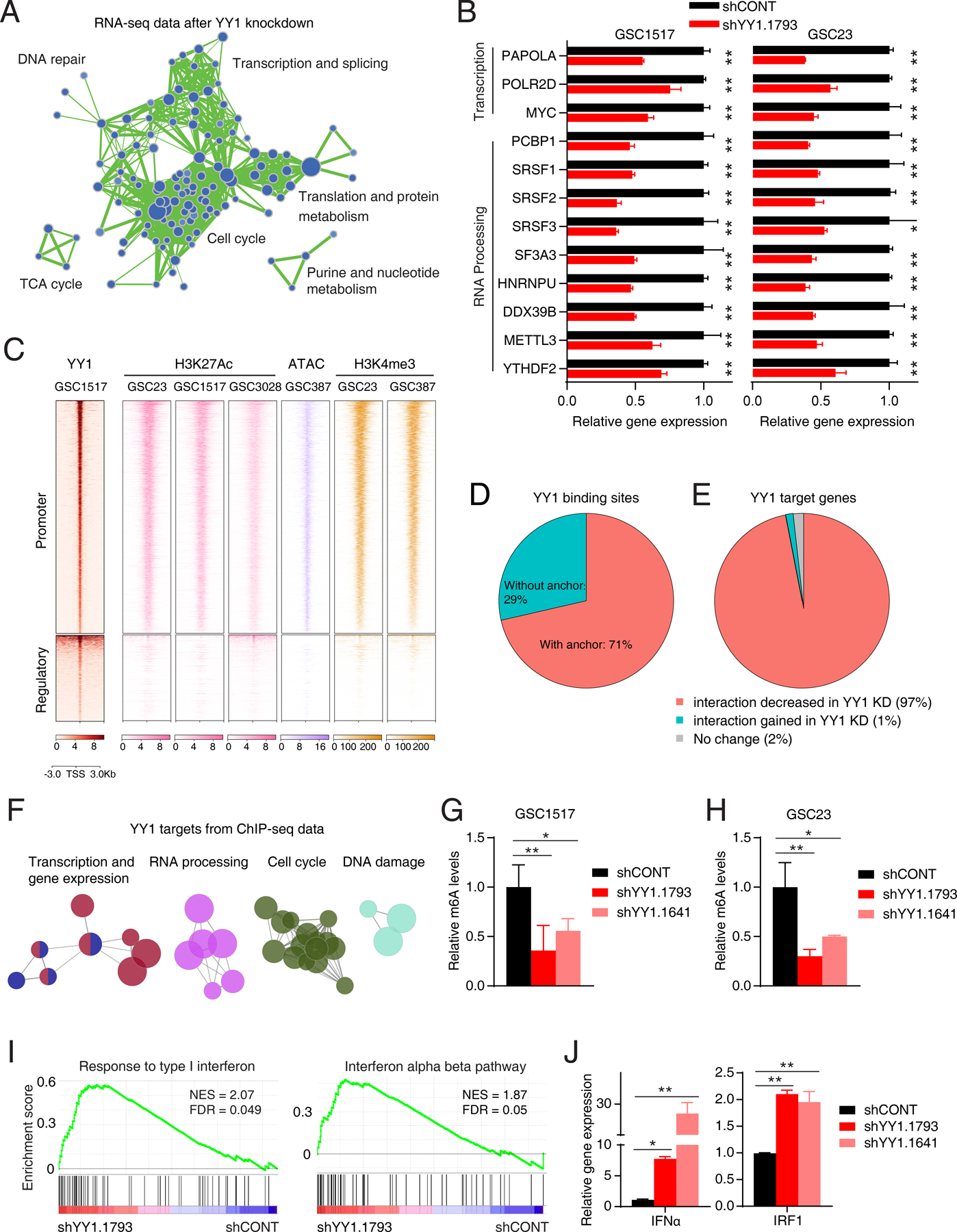
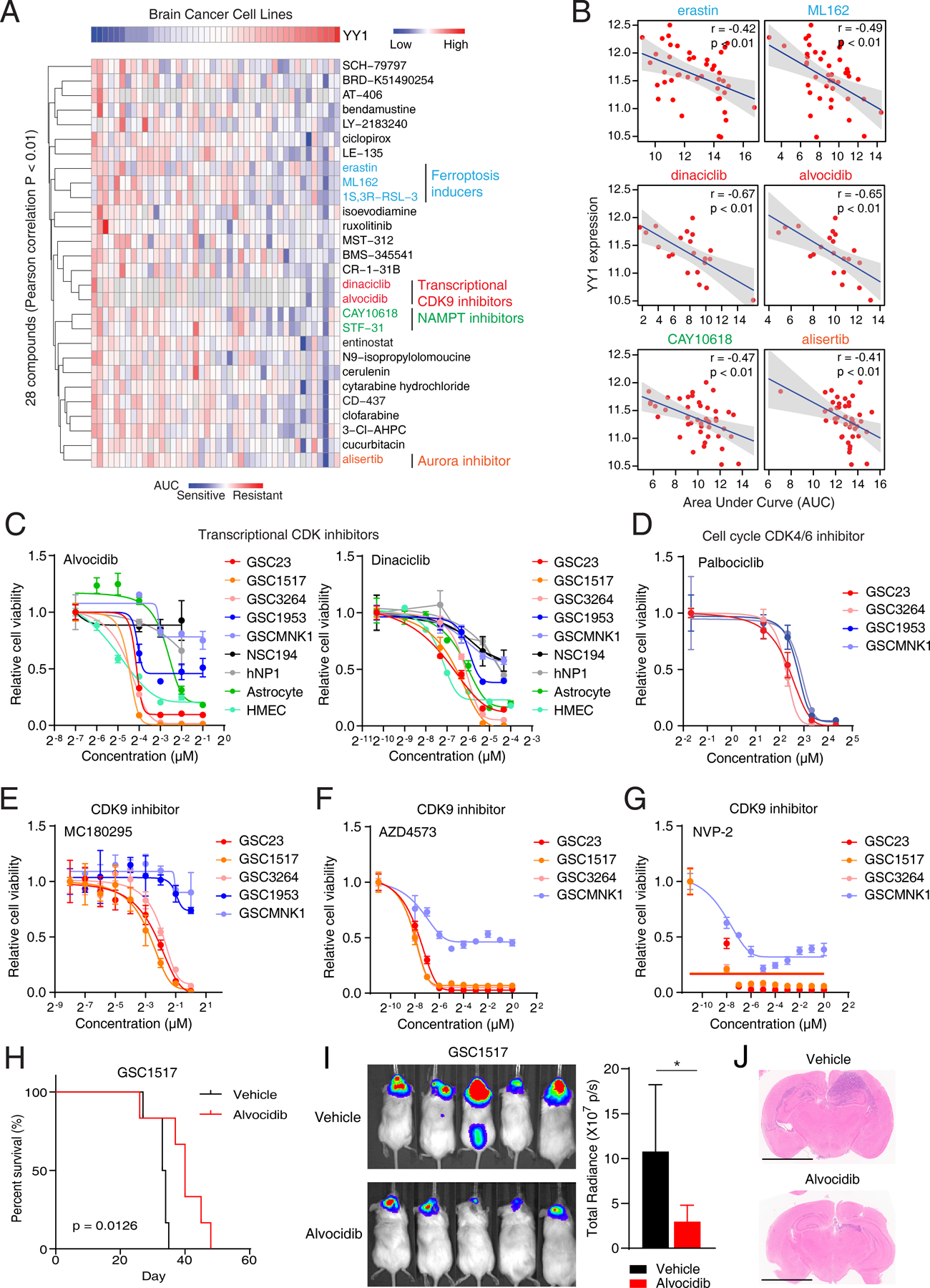
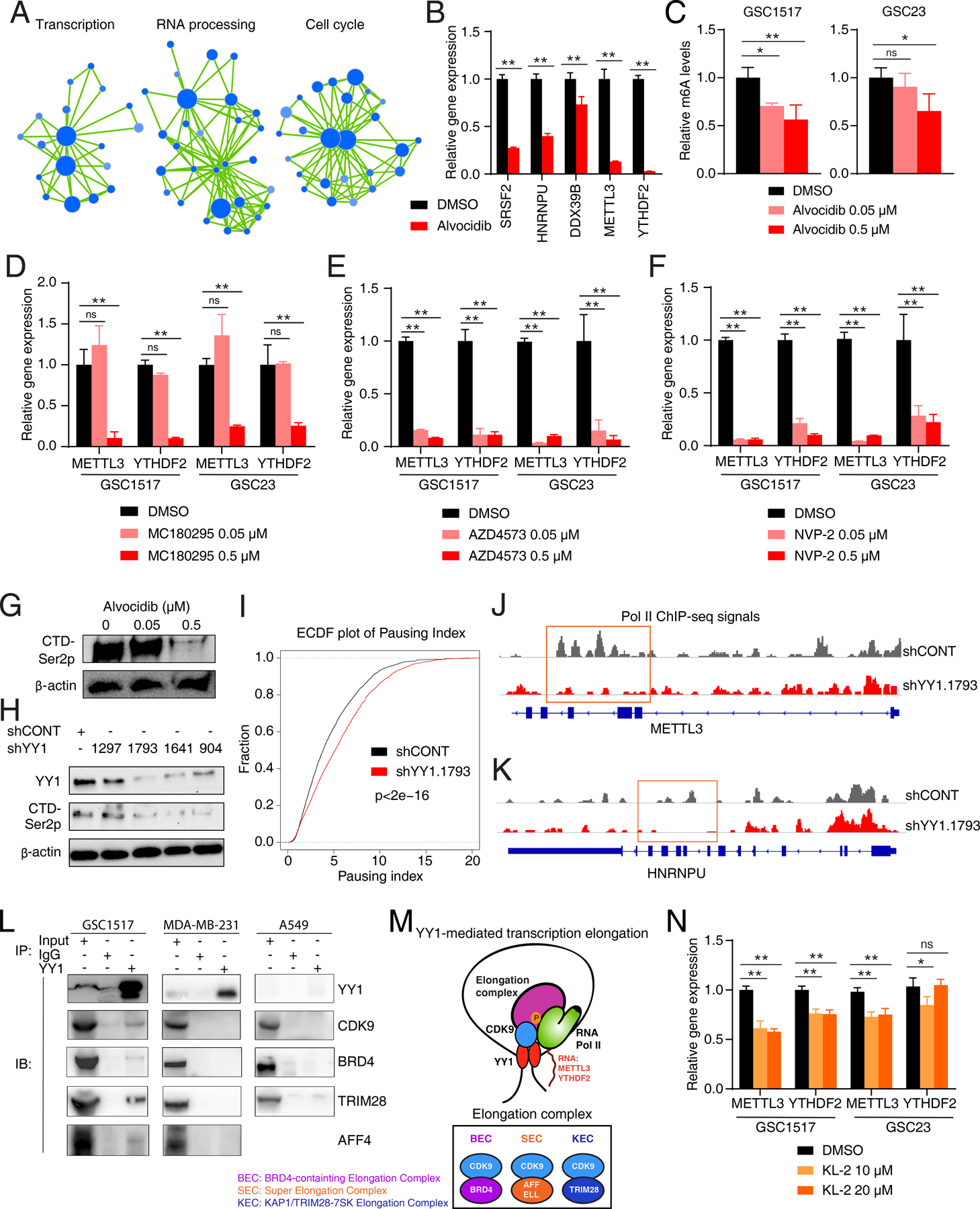
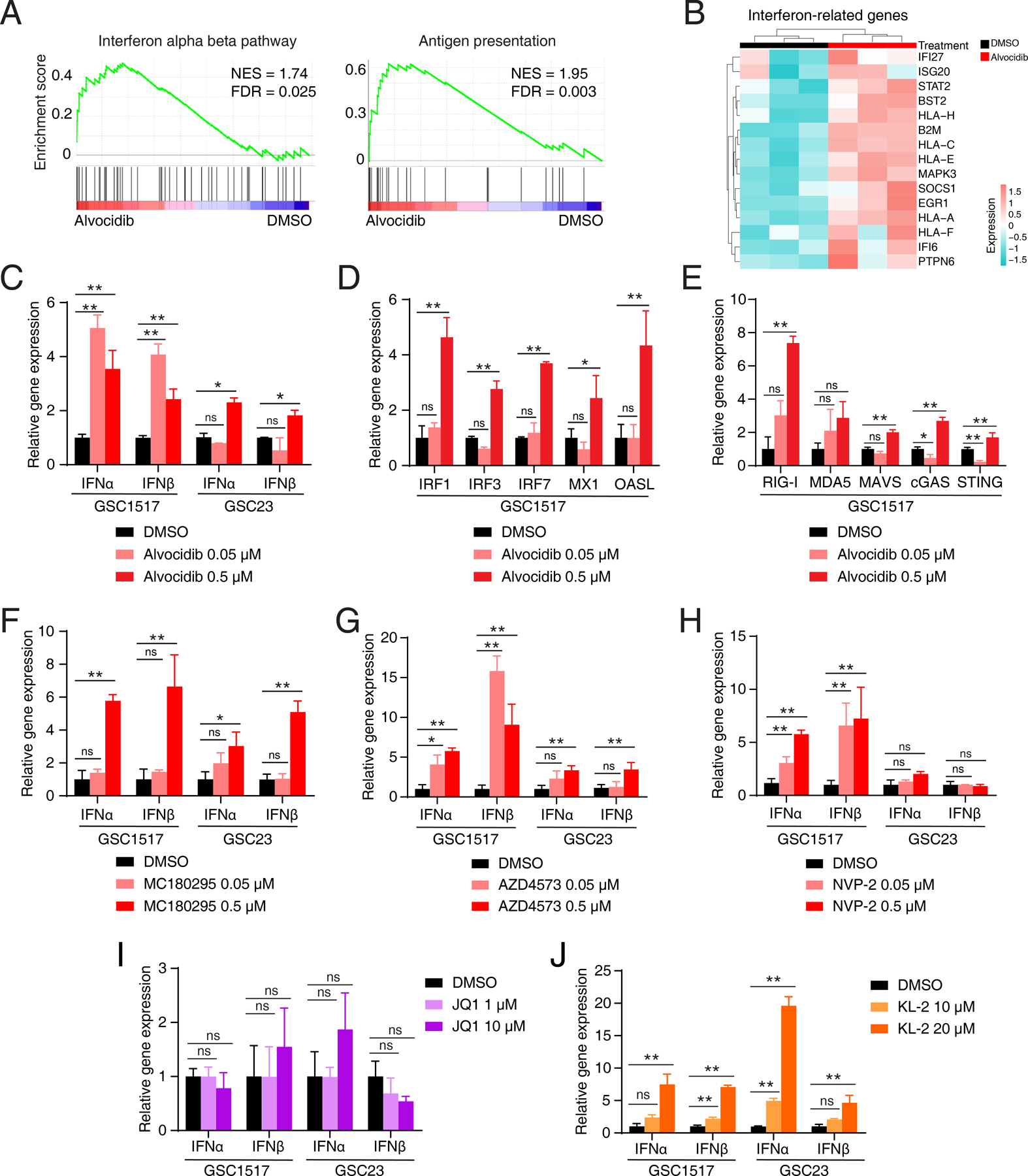
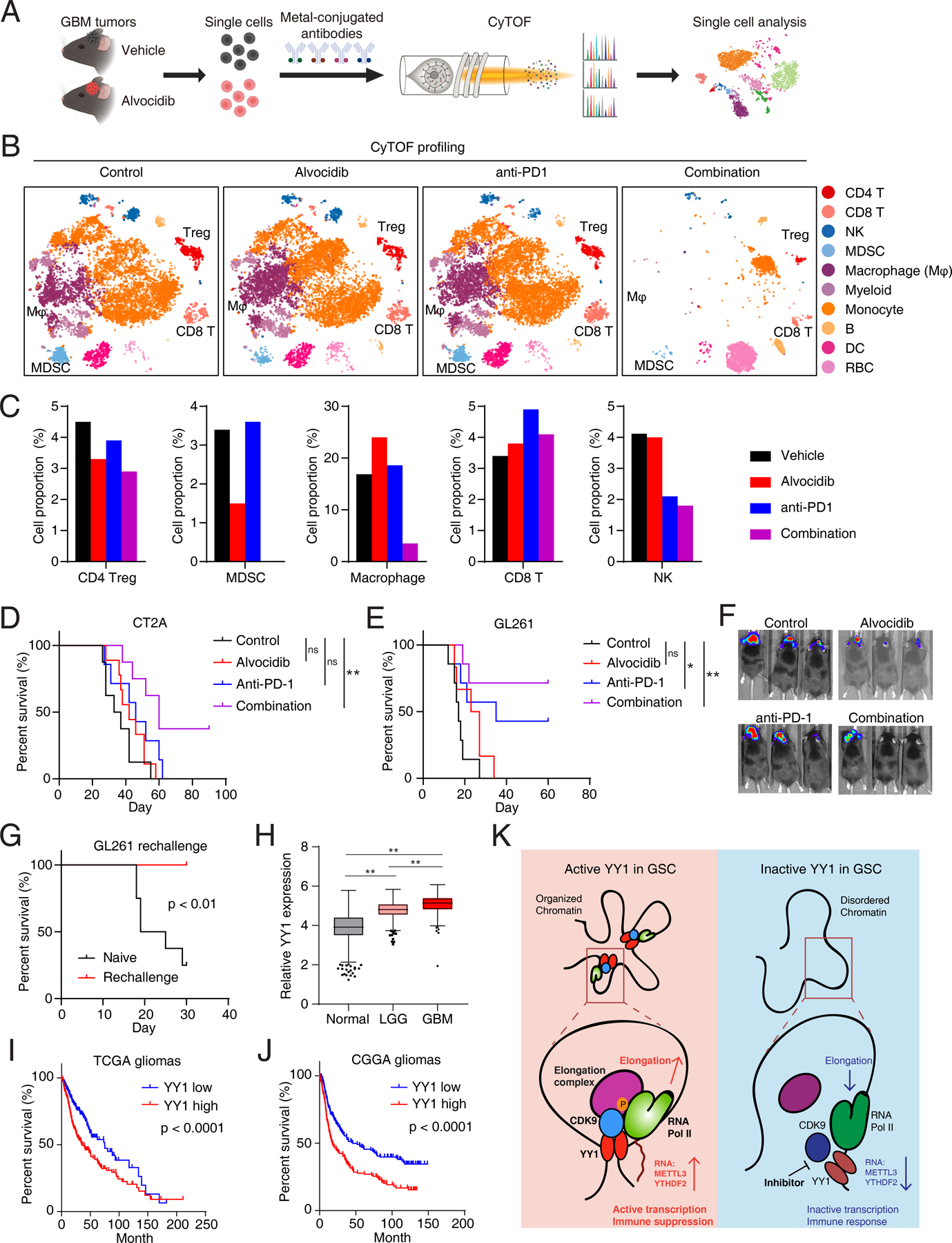
Glioblastoma (GBM) is the most lethal primary brain cancer characterized by therapeutic resistance, which is promoted by GBM stem cells (GSC). Here, we interrogated gene expression and whole-genome CRISPR/Cas9 screening in a large panel of patient-derived GSCs, differentiated GBM cells (DGC), and neural stem cells (NSC) to identify master regulators of GSC stemness, revealing an essential transcription state with increased RNA polymerase II-mediated transcription. The YY1 and transcriptional CDK9 complex was essential for GSC survival and maintenance in vitro and in vivo. YY1 interacted with CDK9 to regulate transcription elongation in GSCs. Genetic or pharmacologic targeting of the YY1-CDK9 complex elicited RNA m6A modification-dependent interferon responses, reduced regulatory T-cell infiltration, and augmented efficacy of immune checkpoint therapy in GBM. Collectively, these results suggest that YY1-CDK9 transcription elongation complex defines a targetable cell state with active transcription, suppressed interferon responses, and immunotherapy resistance in GBM. SIGNIFICANCE: Effective strategies to rewire immunosuppressive microenvironment and enhance immunotherapy response are still lacking in GBM. YY1-driven transcriptional elongation machinery represents a druggable target to activate interferon response and enhance anti-PD-1 response through regulating the m6A modification program, linking epigenetic regulation to immunomodulatory function in GBM.This article is highlighted in the In This Issue feature, p. 275.
©2021 American Association for Cancer Research.
Figures
References
-
- Singh SK, Hawkins C, Clarke ID, Squire JA, Bayani J, Hide T, et al. Identification of human brain tumour initiating cells. Nature 2004;432(7015):396–401 - PubMed
Publication types
MeSH terms
Grants and funding
- T32 CA094186/CA/NCI NIH HHS/United States
- F30 CA217065/CA/NCI NIH HHS/United States
- P30 CA030199/CA/NCI NIH HHS/United States
- R01 AI151123/AI/NIAID NIH HHS/United States
- F30 CA217066/CA/NCI NIH HHS/United States
- R01 NS103434/NS/NINDS NIH HHS/United States
- P30 CA014195/CA/NCI NIH HHS/United States
- P30 CA023100/CA/NCI NIH HHS/United States
- R01 CA238662/CA/NCI NIH HHS/United States
- R01 AI107027/AI/NIAID NIH HHS/United States
- R35 CA197718/CA/NCI NIH HHS/United States
- T32 GM007250/GM/NIGMS NIH HHS/United States
- S10 OD018499/OD/NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
