

Genetic dissection of the glutamatergic neuron system in cerebral cortex
- PMID: 34616069
- PMCID: PMC8494647
- DOI: 10.1038/s41586-021-03955-9
Genetic dissection of the glutamatergic neuron system in cerebral cortex
Abstract
Diverse types of glutamatergic pyramidal neurons mediate the myriad processing streams and output channels of the cerebral cortex1,2, yet all derive from neural progenitors of the embryonic dorsal telencephalon3,4. Here we establish genetic strategies and tools for dissecting and fate-mapping subpopulations of pyramidal neurons on the basis of their developmental and molecular programs. We leverage key transcription factors and effector genes to systematically target temporal patterning programs in progenitors and differentiation programs in postmitotic neurons. We generated over a dozen temporally inducible mouse Cre and Flp knock-in driver lines to enable the combinatorial targeting of major progenitor types and projection classes. Combinatorial strategies confer viral access to subsets of pyramidal neurons defined by developmental origin, marker expression, anatomical location and projection targets. These strategies establish an experimental framework for understanding the hierarchical organization and developmental trajectory of subpopulations of pyramidal neurons that assemble cortical processing networks and output channels.
© 2021. The Author(s).
Conflict of interest statement
J.A.H. is currently employed by Cajal Neuroscience. The remaining authors declare no competing interests.
Figures


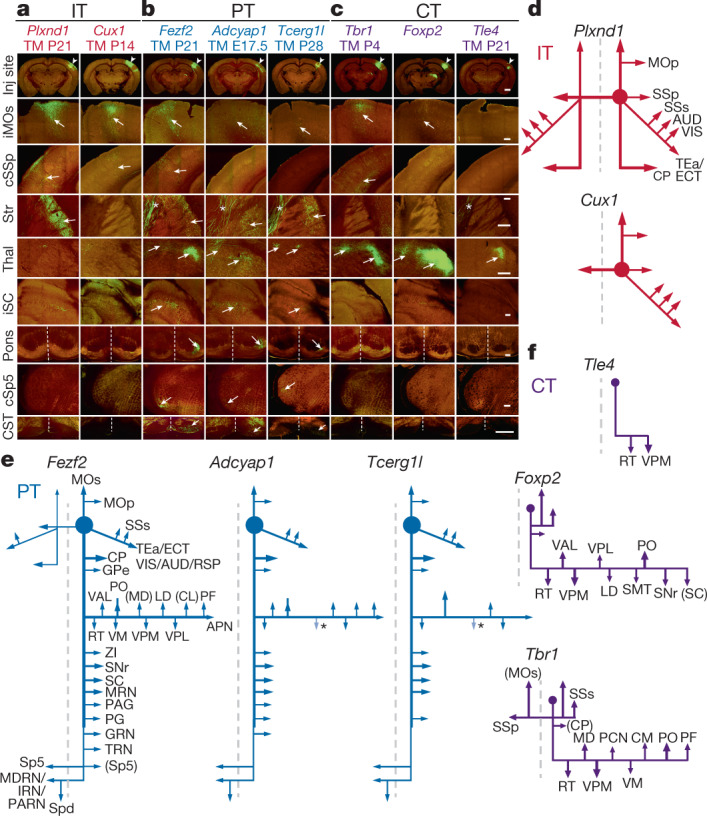




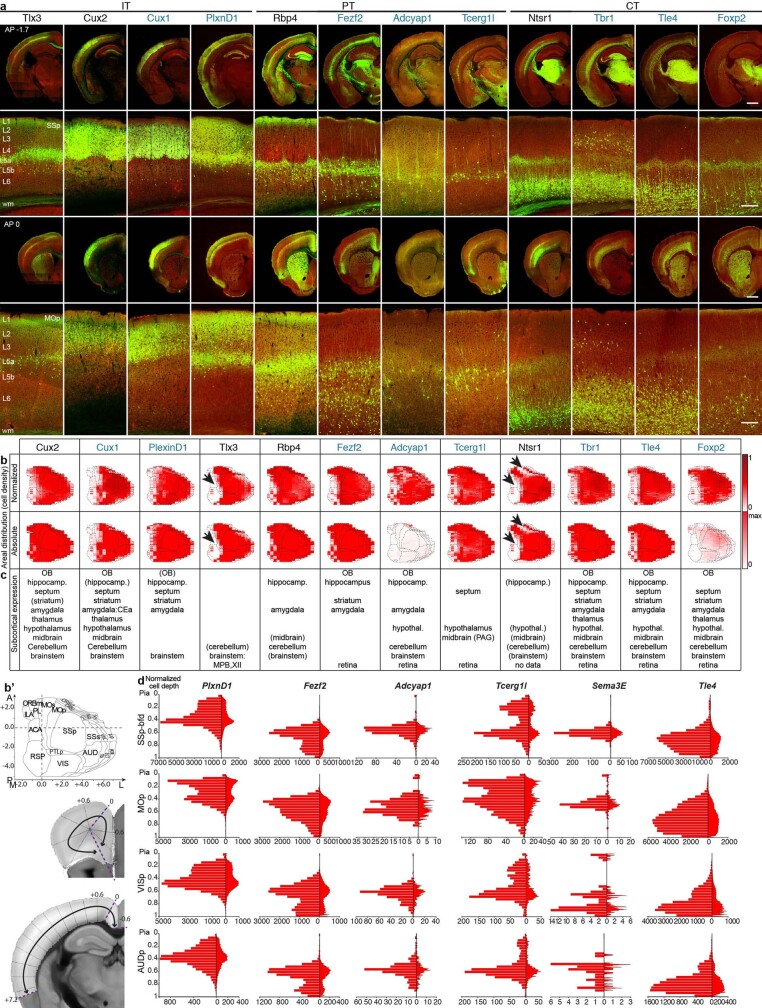




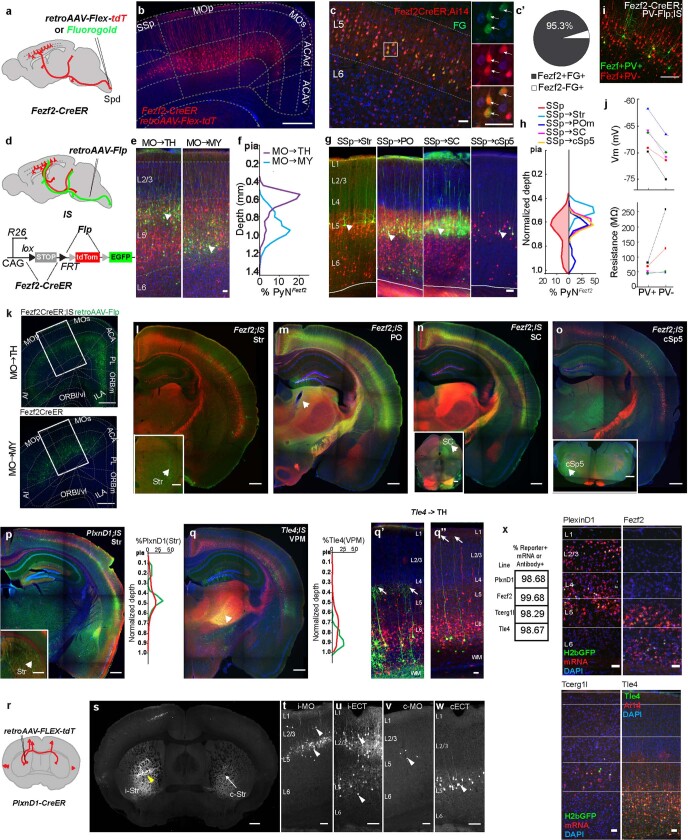

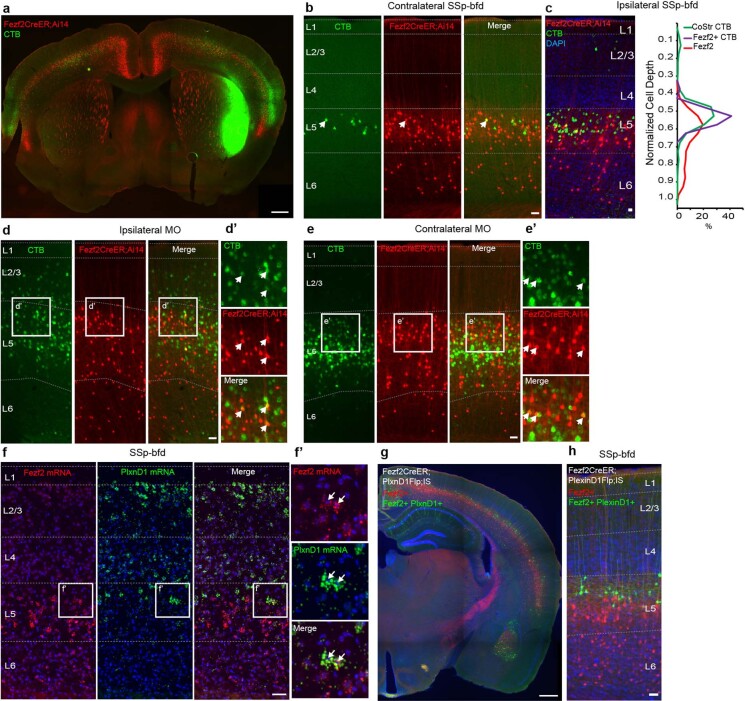
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
