

Cellular anatomy of the mouse primary motor cortex
- PMID: 34616071
- PMCID: PMC8494646
- DOI: 10.1038/s41586-021-03970-w
Cellular anatomy of the mouse primary motor cortex
Abstract
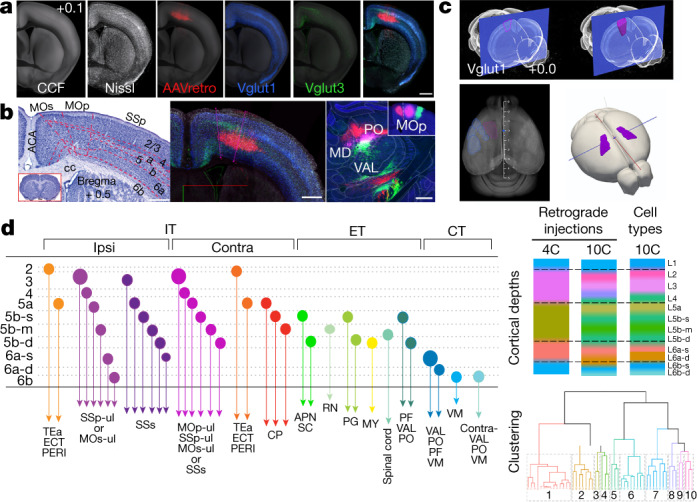
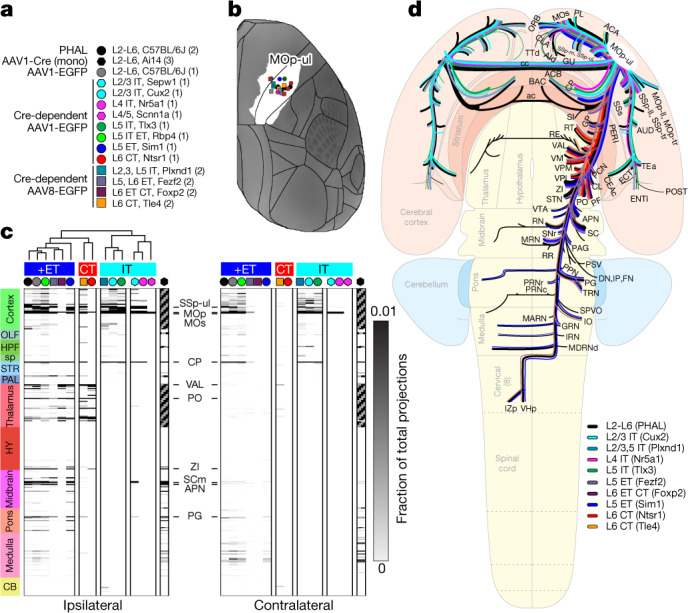
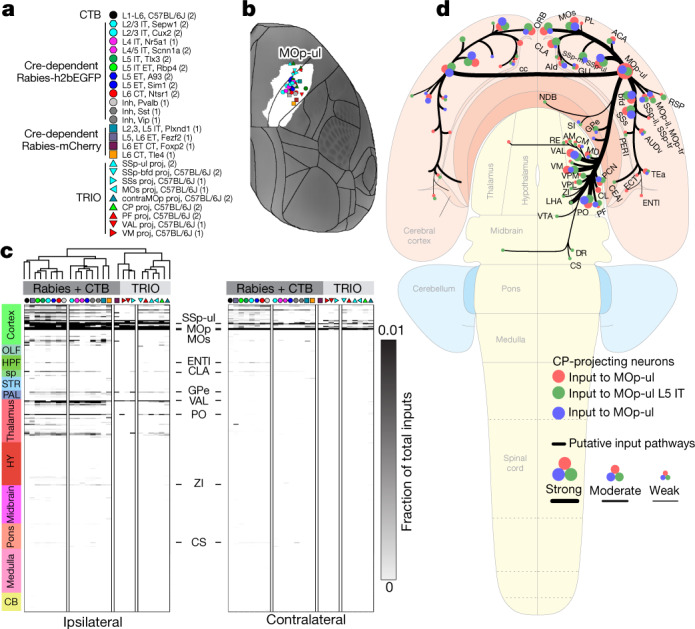
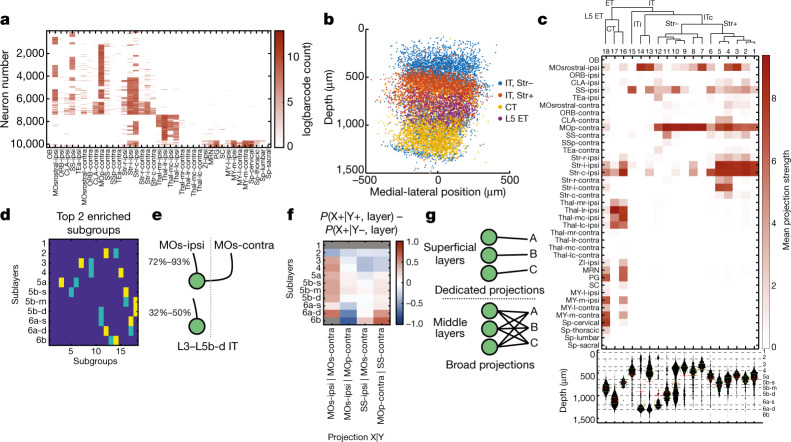
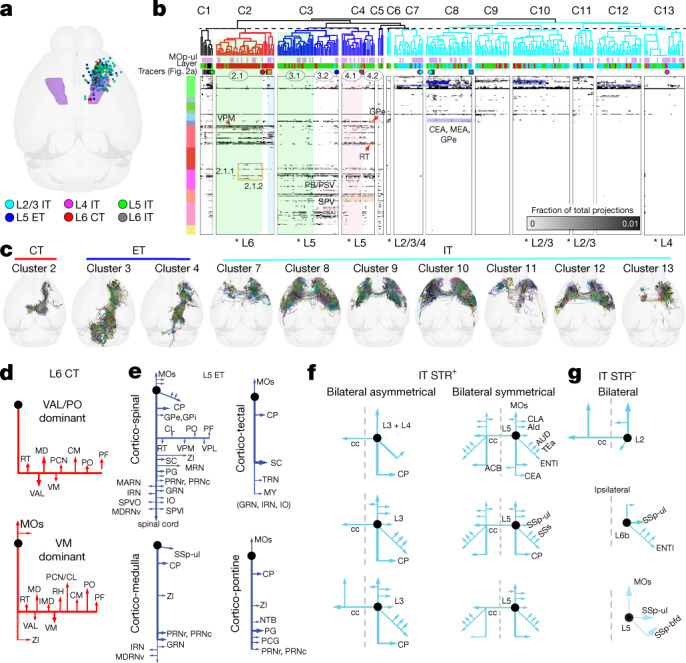
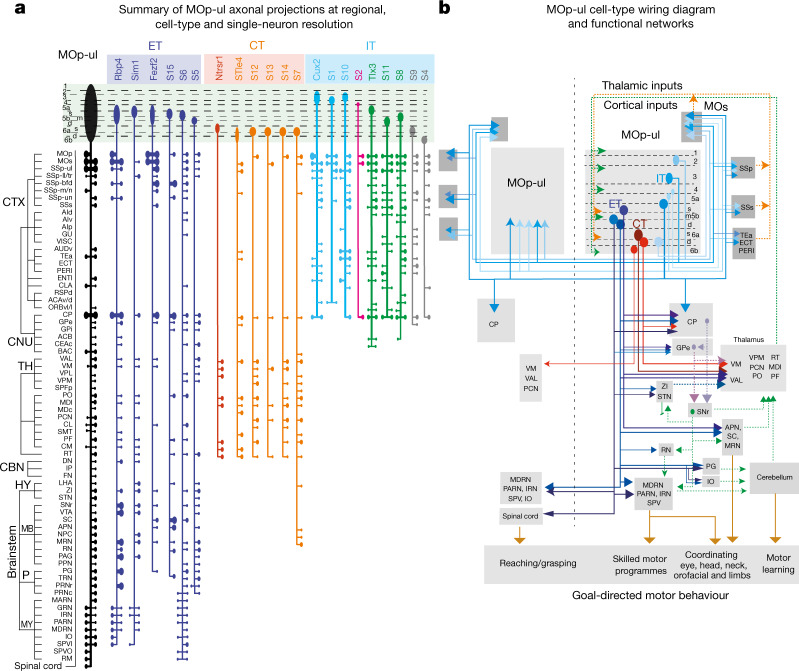
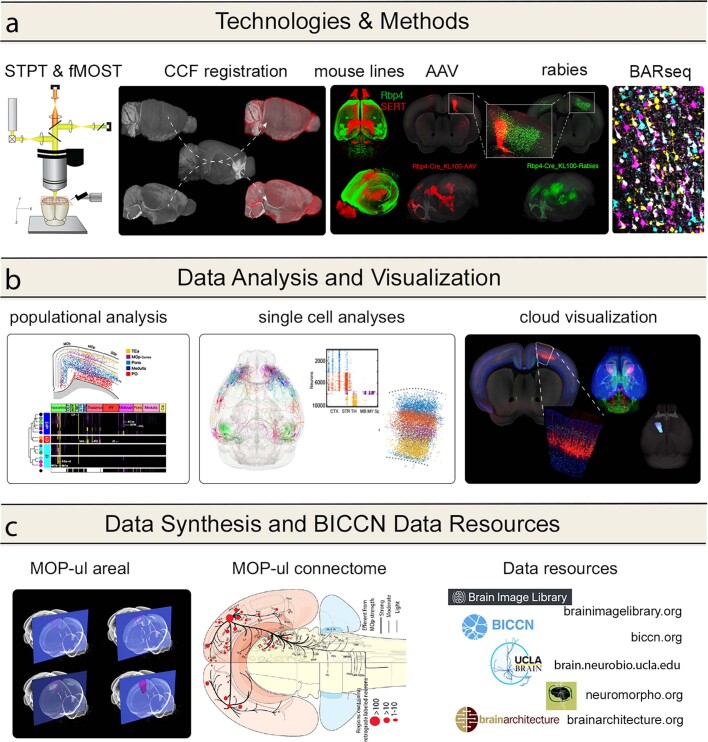
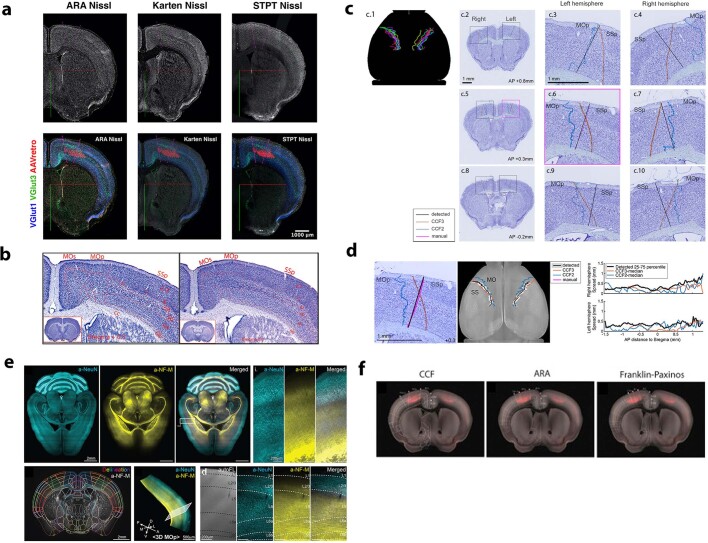
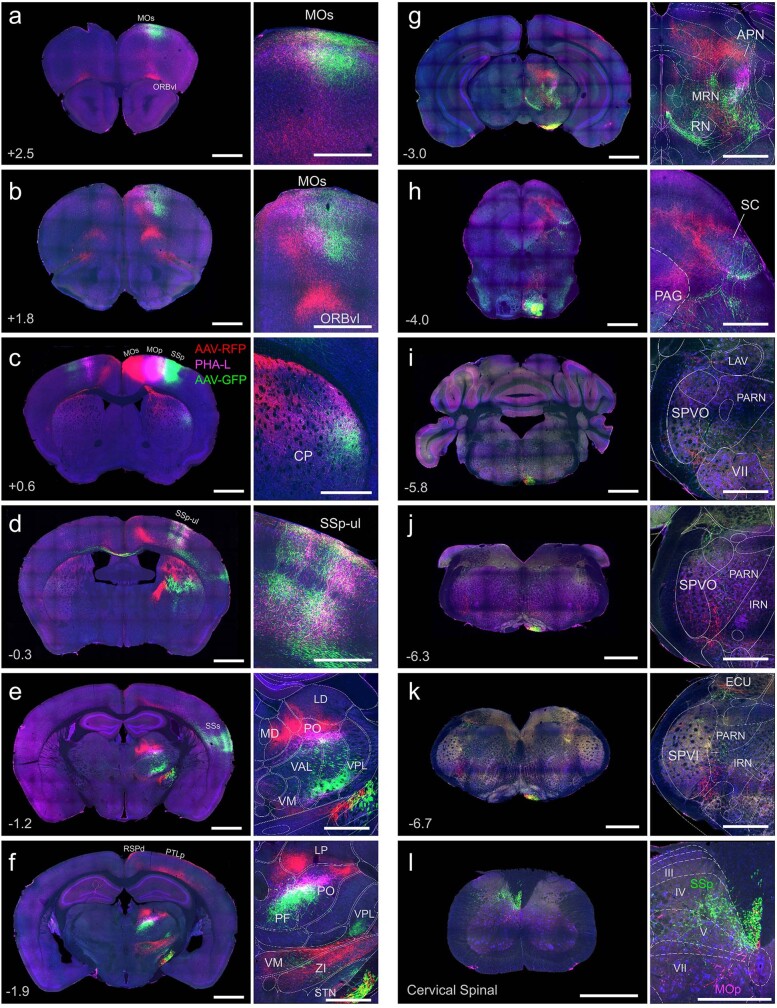
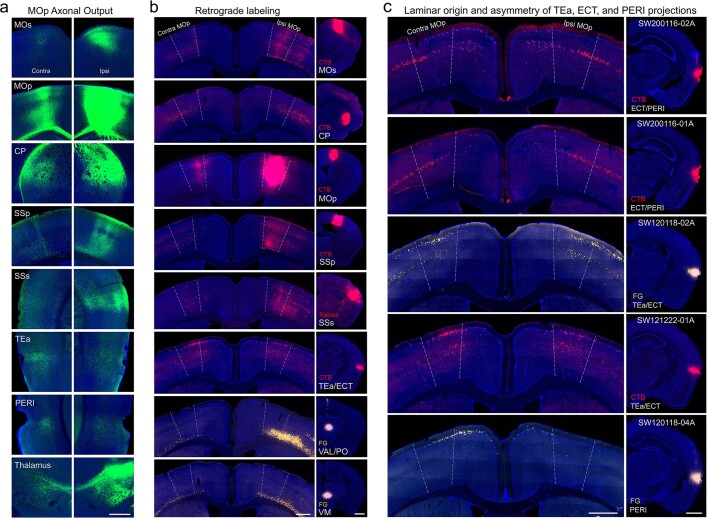
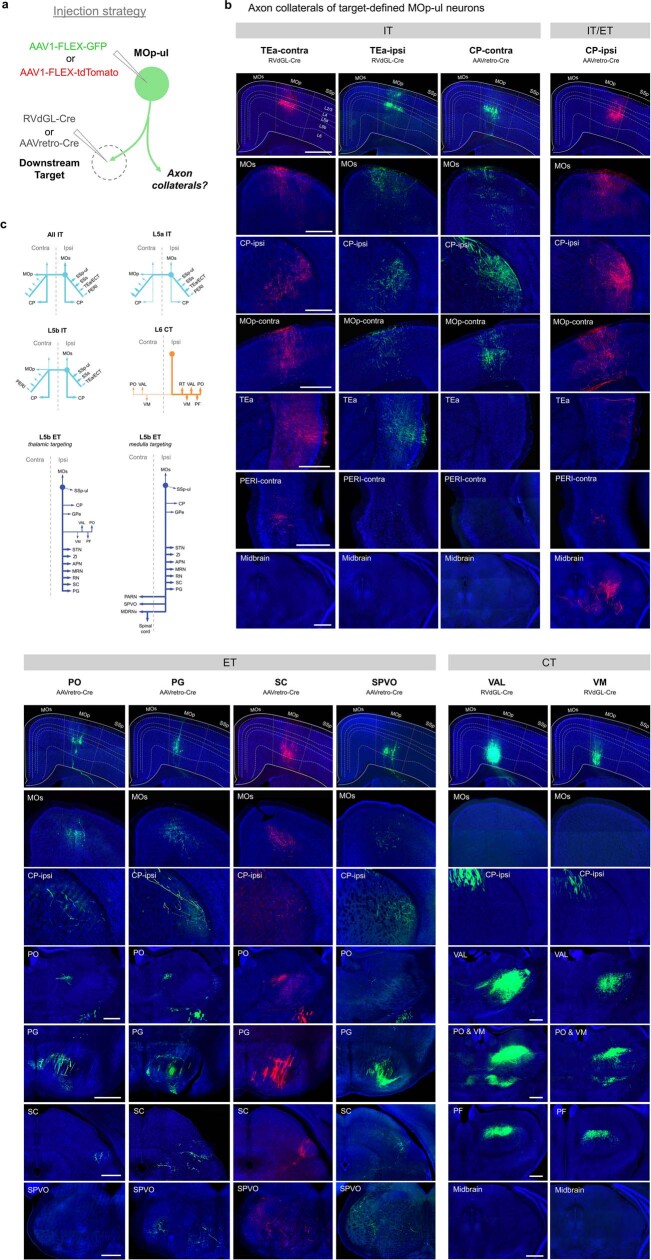
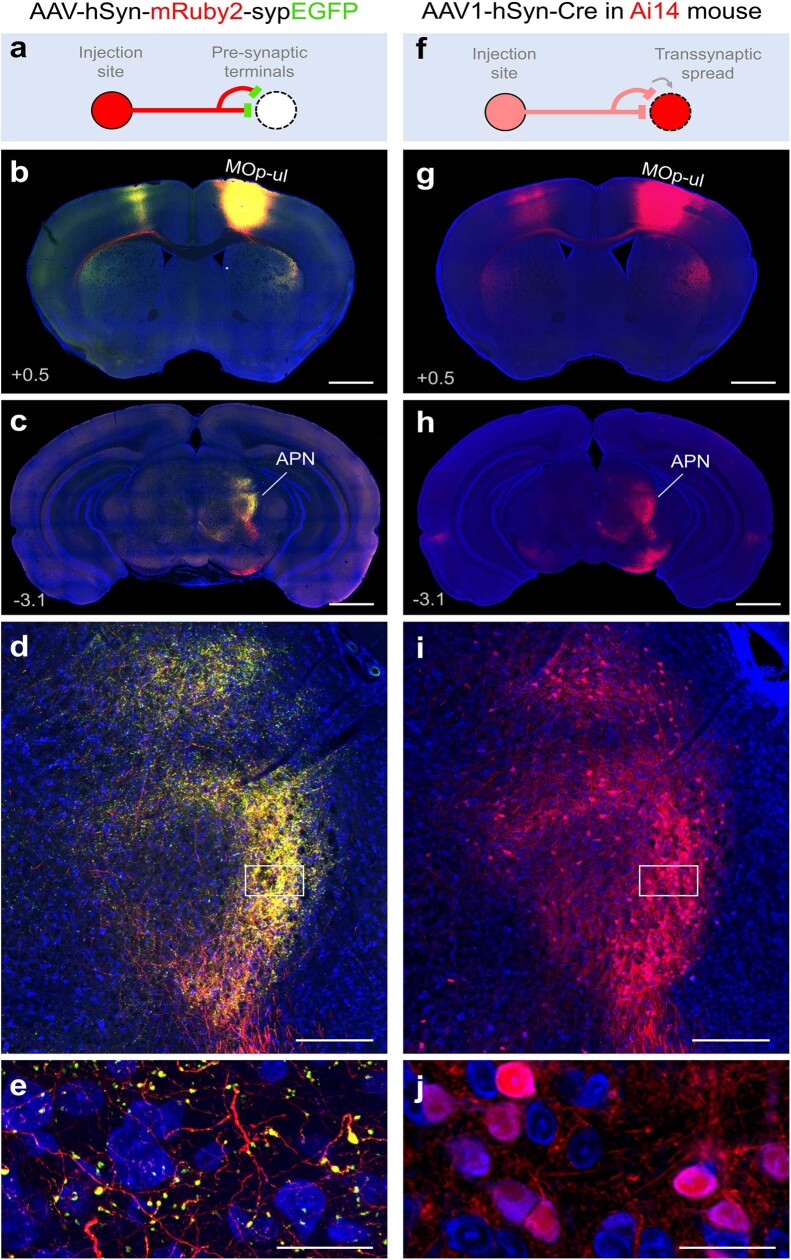
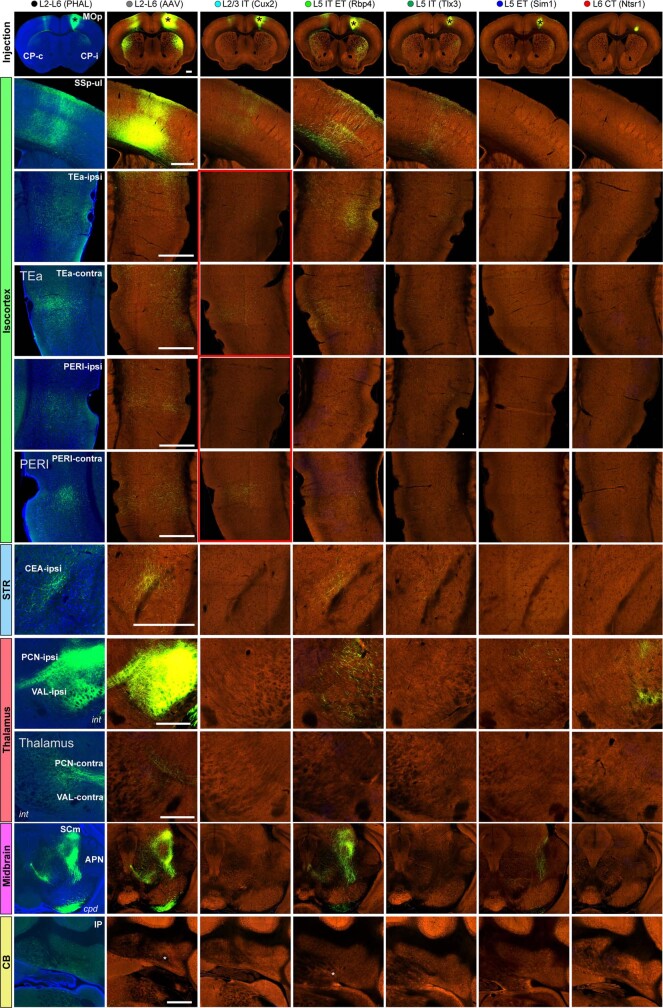
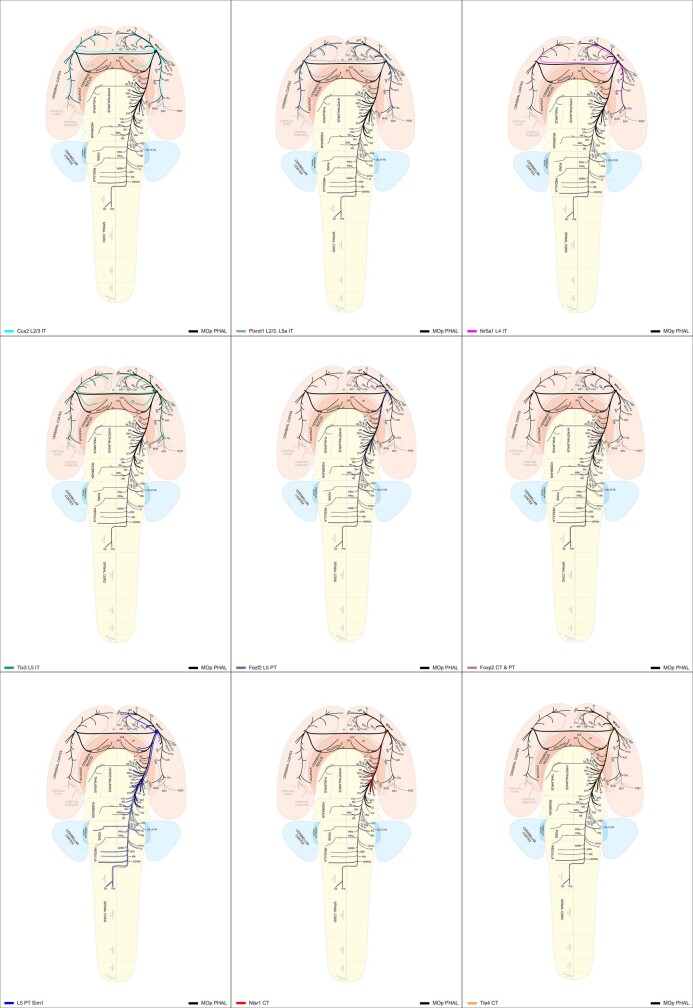
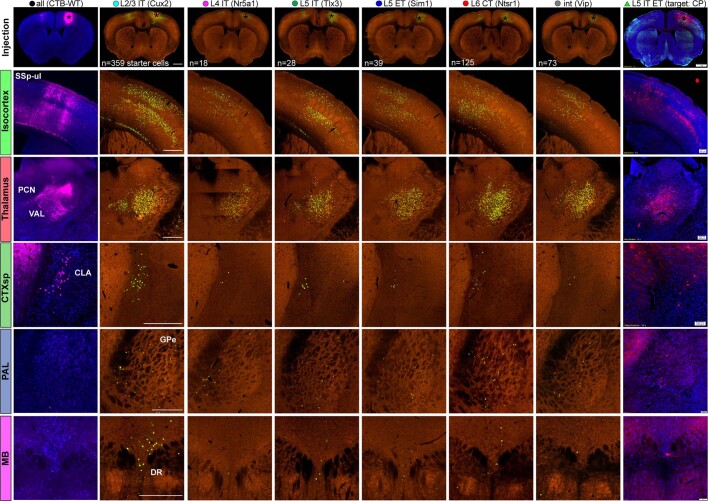
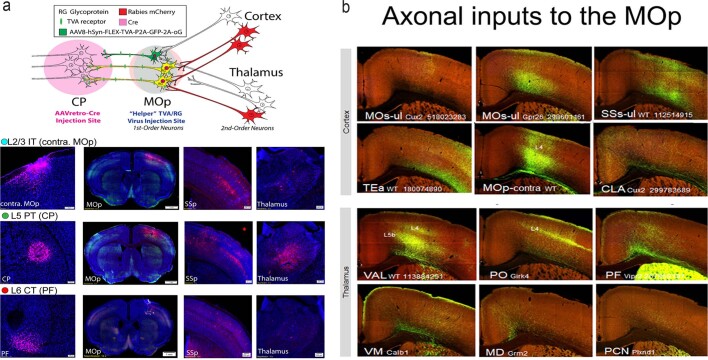
An essential step toward understanding brain function is to establish a structural framework with cellular resolution on which multi-scale datasets spanning molecules, cells, circuits and systems can be integrated and interpreted1. Here, as part of the collaborative Brain Initiative Cell Census Network (BICCN), we derive a comprehensive cell type-based anatomical description of one exemplar brain structure, the mouse primary motor cortex, upper limb area (MOp-ul). Using genetic and viral labelling, barcoded anatomy resolved by sequencing, single-neuron reconstruction, whole-brain imaging and cloud-based neuroinformatics tools, we delineated the MOp-ul in 3D and refined its sublaminar organization. We defined around two dozen projection neuron types in the MOp-ul and derived an input-output wiring diagram, which will facilitate future analyses of motor control circuitry across molecular, cellular and system levels. This work provides a roadmap towards a comprehensive cellular-resolution description of mammalian brain architecture.
© 2021. The Author(s).
Conflict of interest statement
A.M.Z. is a founder and equity owner of Cajal Neuroscience and a member of its scientific advisory board. J.A.H., K.E.H. and P.R.N. are currently employed by Cajal Neuroscience.
Figures






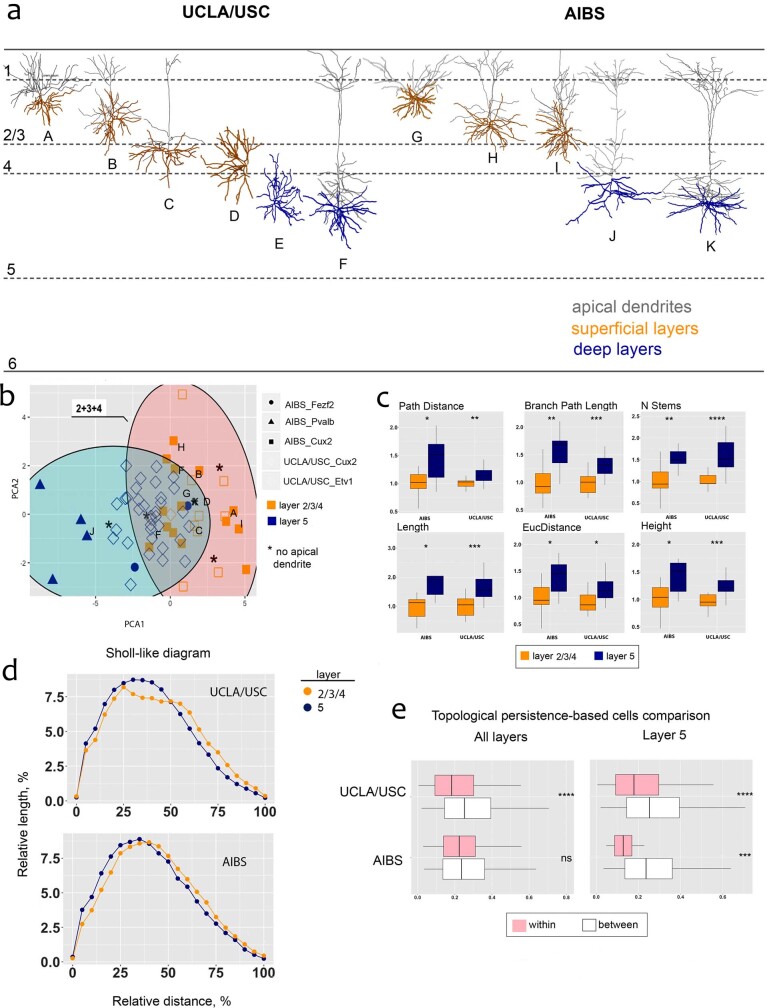

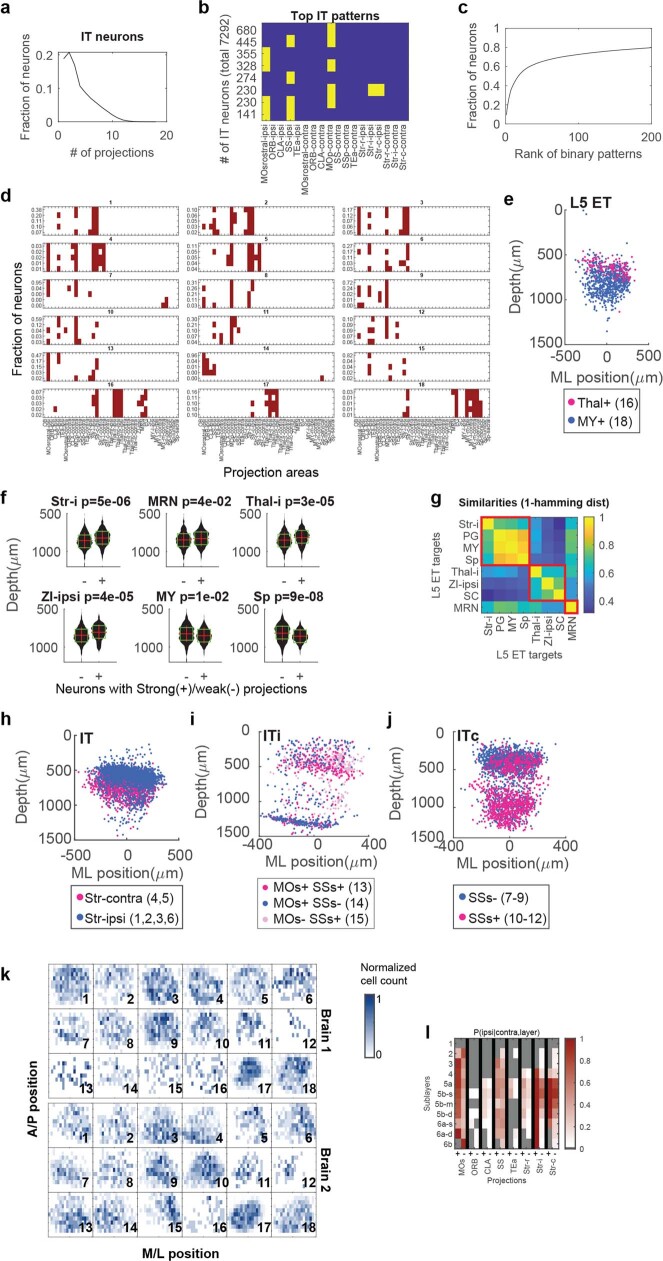
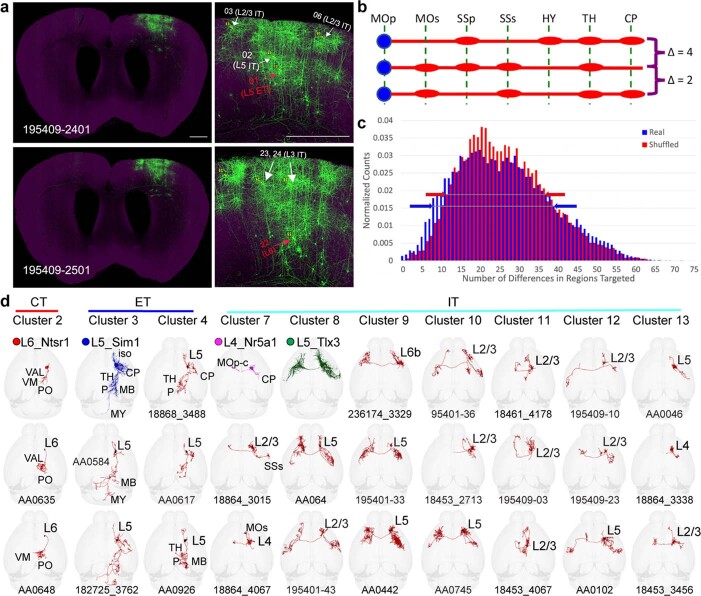

References
Publication types
MeSH terms
Substances
Grants and funding
- P41 EB015922/EB/NIBIB NIH HHS/United States
- U01 MH106008/MH/NIMH NIH HHS/United States
- U19 MH114821/MH/NIMH NIH HHS/United States
- EY019049/NH/NIH HHS/United States
- R01 EB022899/EB/NIBIB NIH HHS/United States
- R01 DC008983/DC/NIDCD NIH HHS/United States
- MH114821/NH/NIH HHS/United States
- RF1 MH124605/MH/NIMH NIH HHS/United States
- R01 EY023173/EY/NEI NIH HHS/United States
- RF1 AG064049/AG/NIA NIH HHS/United States
- RF1 MH114112/MH/NIMH NIH HHS/United States
- R01 NS073129/NS/NINDS NIH HHS/United States
- U19 MH114830/MH/NIMH NIH HHS/United States
- R01 NS086082/NS/NINDS NIH HHS/United States
- R01 MH094360/MH/NIMH NIH HHS/United States
- 5R01 NS073129/NH/NIH HHS/United States
- U01 MH114829/MH/NIMH NIH HHS/United States
- MH114824/NH/NIH HHS/United States
- U01 MH114824/MH/NIMH NIH HHS/United States
- R01 NS096720/NS/NINDS NIH HHS/United States
- R01 MH116176/MH/NIMH NIH HHS/United States
- NS107466/NH/NIH HHS/United States
- R01 MH113005/MH/NIMH NIH HHS/United States
- EB022899/NH/NIH HHS/United States
- R01 EY019049/EY/NEI NIH HHS/United States
- U01 MH109113/MH/NIMH NIH HHS/United States
- RF1 MH114132/MH/NIMH NIH HHS/United States
- U01 MH117079/MH/NIMH NIH HHS/United States
- 5U19 MH114821-03/NH/NIH HHS/United States
- R01 DA036913/DA/NIDA NIH HHS/United States
- U01 MH105982/MH/NIMH NIH HHS/United States
- U19 MH114831/MH/NIMH NIH HHS/United States
- R01 LM012736/LM/NLM NIH HHS/United States
- RF1 MH12460501/NH/NIH HHS/United States
- R01 NS039600/NS/NINDS NIH HHS/United States
- 5R01 DA036913/NH/NIH HHS/United States
- U01 MH116990/MH/NIMH NIH HHS/United States
- U19 NS107466/NS/NINDS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
