

Otic Neurogenesis in Xenopus laevis: Proliferation, Differentiation, and the Role of Eya1
- PMID: 34616280
- PMCID: PMC8488300
- DOI: 10.3389/fnana.2021.722374
Otic Neurogenesis in Xenopus laevis: Proliferation, Differentiation, and the Role of Eya1
Abstract
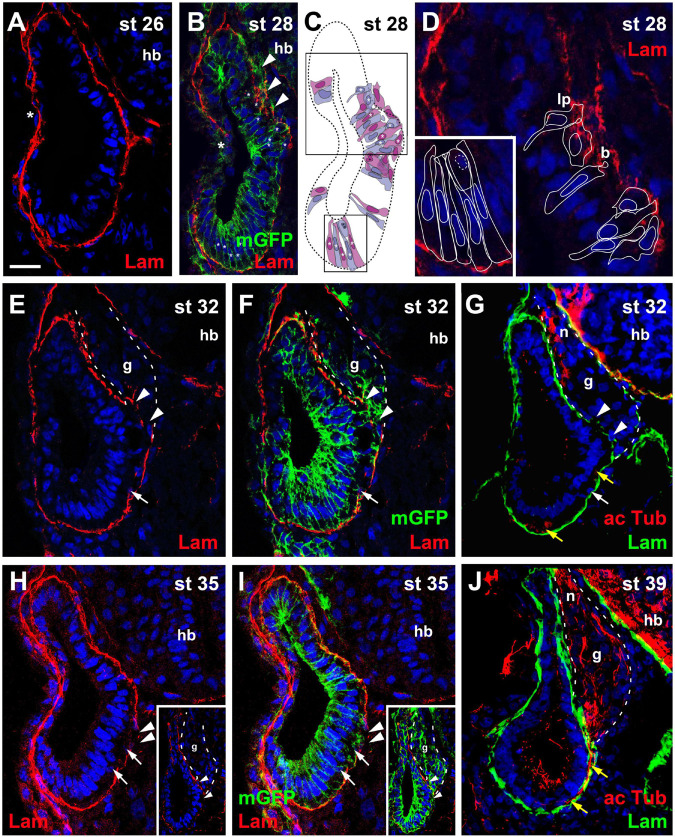
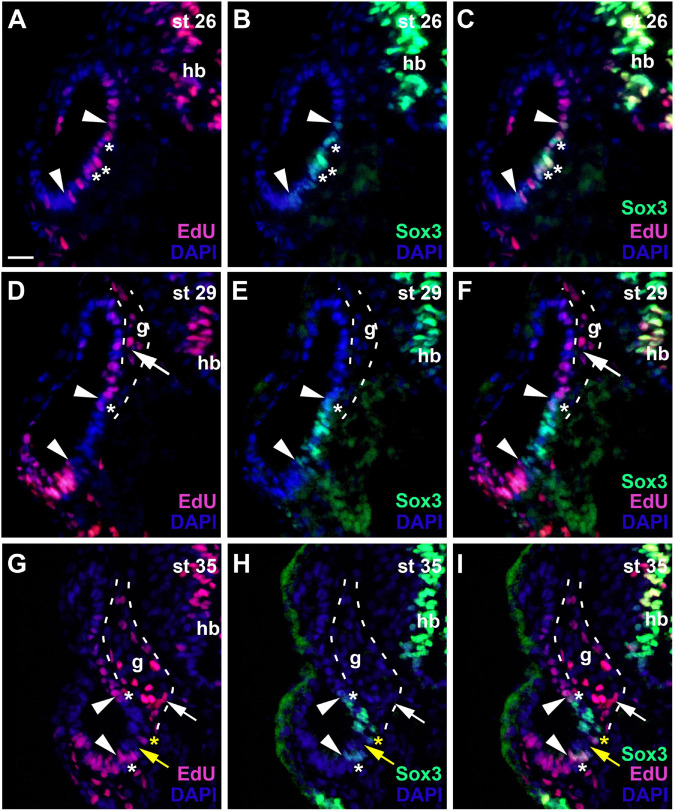
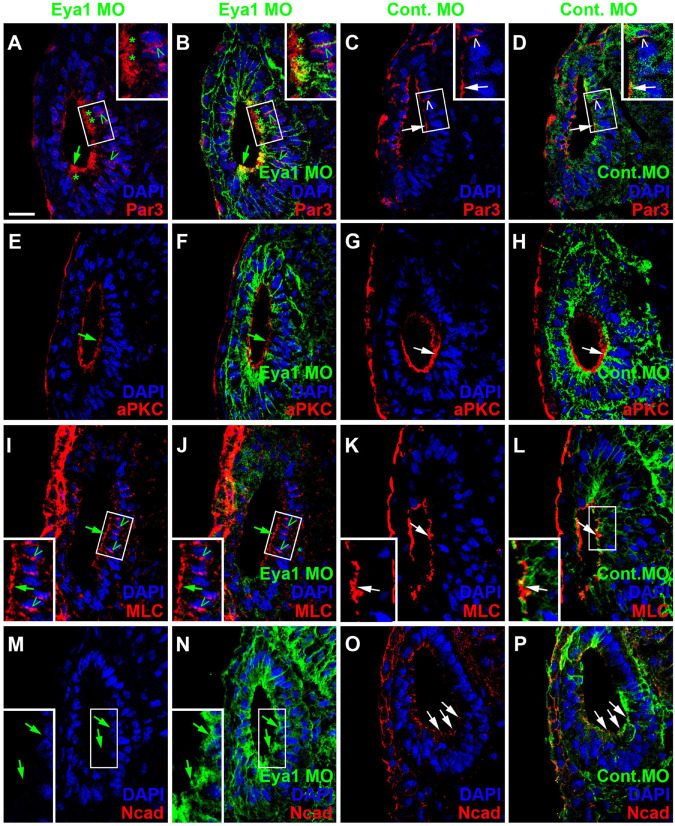
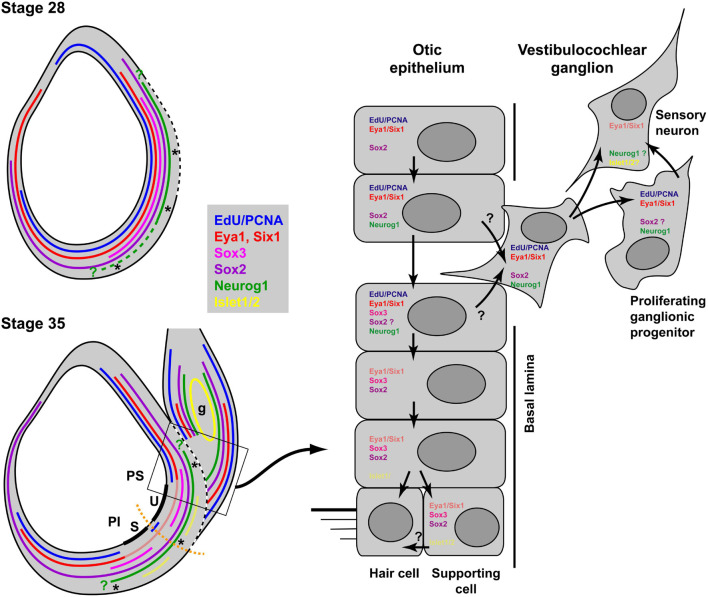
Using immunostaining and confocal microscopy, we here provide the first detailed description of otic neurogenesis in Xenopus laevis. We show that the otic vesicle comprises a pseudostratified epithelium with apicobasal polarity (apical enrichment of Par3, aPKC, phosphorylated Myosin light chain, N-cadherin) and interkinetic nuclear migration (apical localization of mitotic, pH3-positive cells). A Sox3-immunopositive neurosensory area in the ventromedial otic vesicle gives rise to neuroblasts, which delaminate through breaches in the basal lamina between stages 26/27 and 39. Delaminated cells congregate to form the vestibulocochlear ganglion, whose peripheral cells continue to proliferate (as judged by EdU incorporation), while central cells differentiate into Islet1/2-immunopositive neurons from stage 29 on and send out neurites at stage 31. The central part of the neurosensory area retains Sox3 but stops proliferating from stage 33, forming the first sensory areas (utricular/saccular maculae). The phosphatase and transcriptional coactivator Eya1 has previously been shown to play a central role for otic neurogenesis but the underlying mechanism is poorly understood. Using an antibody specifically raised against Xenopus Eya1, we characterize the subcellular localization of Eya1 proteins, their levels of expression as well as their distribution in relation to progenitor and neuronal differentiation markers during otic neurogenesis. We show that Eya1 protein localizes to both nuclei and cytoplasm in the otic epithelium, with levels of nuclear Eya1 declining in differentiating (Islet1/2+) vestibulocochlear ganglion neurons and in the developing sensory areas. Morpholino-based knockdown of Eya1 leads to reduction of proliferating, Sox3- and Islet1/2-immunopositive cells, redistribution of cell polarity proteins and loss of N-cadherin suggesting that Eya1 is required for maintenance of epithelial cells with apicobasal polarity, progenitor proliferation and neuronal differentiation during otic neurogenesis.
Keywords: Eya1; Xenopus; cell polarity; delamination; ear; neurogenesis; otic vesicle; vestibulocochlear ganglion.
Copyright © 2021 Almasoudi and Schlosser.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures









References
-
- Adam J., Myat A., Leroux I., Eddison M., Henrique D., Ishhorowicz D., et al. (1998). Cell fate choices and the expression of Notch, Delta and Serrate homologues in the chick inner ear: parallels with Drosophila sense-organ development. Development 125 4645–4654. - PubMed
LinkOut - more resources
Full Text Sources
Research Materials