

Physiological roles of Casparian strips and suberin in the transport of water and solutes
- PMID: 34617285
- PMCID: PMC9298204
- DOI: 10.1111/nph.17765
Physiological roles of Casparian strips and suberin in the transport of water and solutes
Abstract
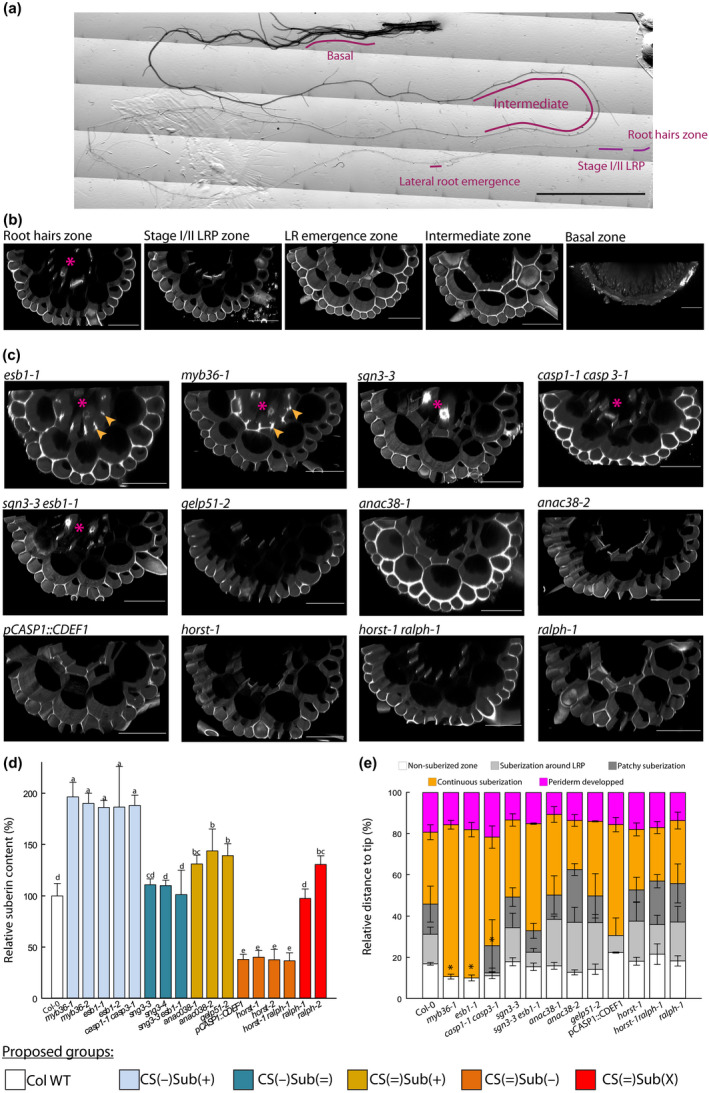
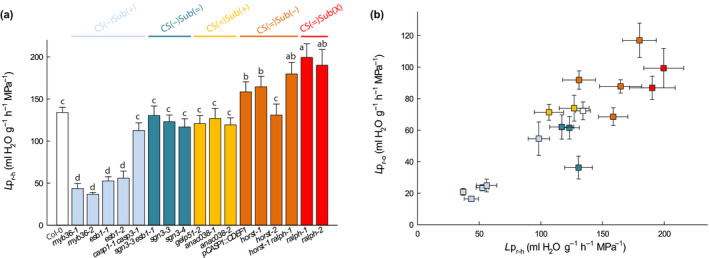
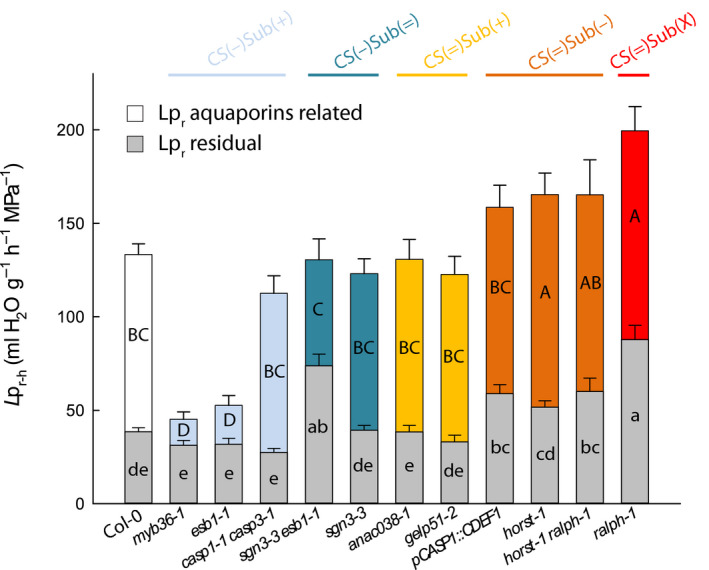
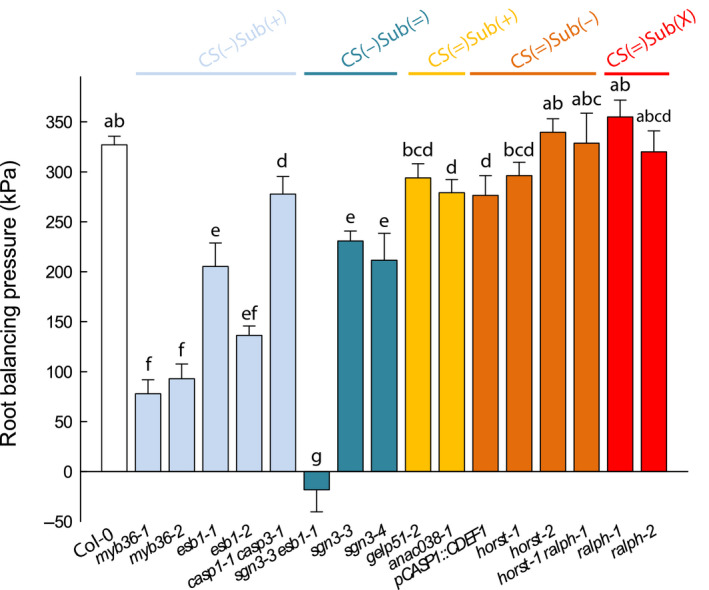
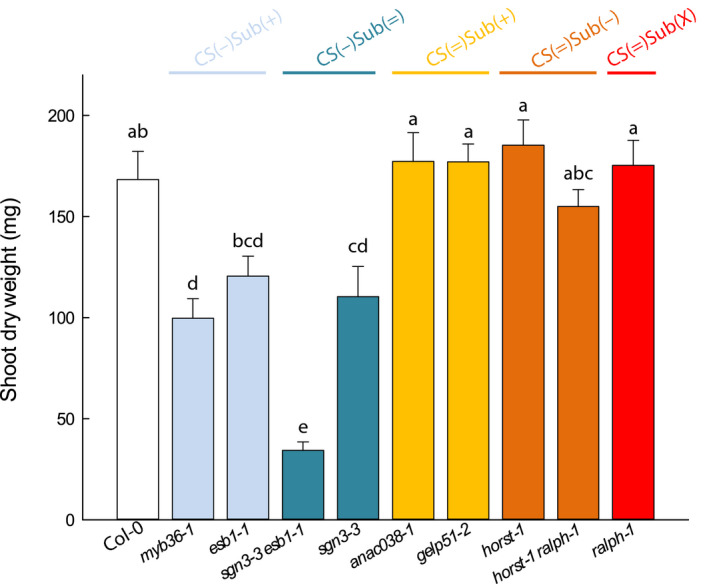
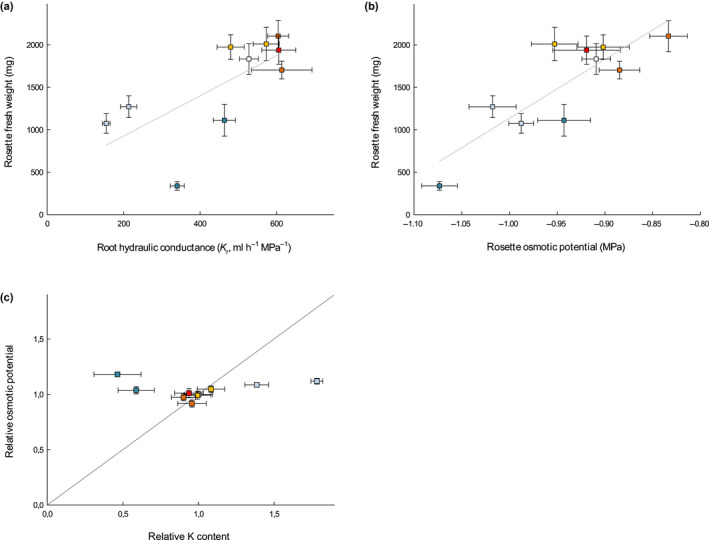
The formation of Casparian strips (CS) and the deposition of suberin at the endodermis of plant roots are thought to limit the apoplastic transport of water and ions. We investigated the specific role of each of these apoplastic barriers in the control of hydro-mineral transport by roots and the consequences on shoot growth. A collection of Arabidopsis thaliana mutants defective in suberin deposition and/or CS development was characterized under standard conditions using a hydroponic system and the Phenopsis platform. Mutants altered in suberin deposition had enhanced root hydraulic conductivity, indicating a restrictive role for this compound in water transport. In contrast, defective CS directly increased solute leakage and indirectly reduced root hydraulic conductivity. Defective CS also led to a reduction in rosette growth, which was partly dependent on the hydro-mineral status of the plant. Ectopic suberin was shown to partially compensate for defective CS phenotypes. Altogether, our work shows that the functionality of the root apoplastic diffusion barriers greatly influences the plant physiology, and that their integrity is tightly surveyed.
Keywords: Arabidopsis thaliana; Casparian strips; apoplastic barriers; aquaporins; root hydraulic conductivity; solutes diffusion; suberin; water transport.
© 2021 The Authors New Phytologist © 2021 New Phytologist Foundation.
Figures

References
-
- Barberon M, Vermeer J, De Bellis D, Wang P, Naseer S, Andersen T, Humbel B, Nawrath C, Takano J, Salt D et al. 2016. Adaptation of root function by nutrient‐induced plasticity of endodermal differentiation. Cell 164: 447–459. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
