

Molecular markers of brain cholesterol homeostasis are unchanged despite a smaller brain mass in a mouse model of cholesteryl ester storage disease
- PMID: 34618372
- PMCID: PMC8766890
- DOI: 10.1002/lipd.12325
Molecular markers of brain cholesterol homeostasis are unchanged despite a smaller brain mass in a mouse model of cholesteryl ester storage disease
Abstract
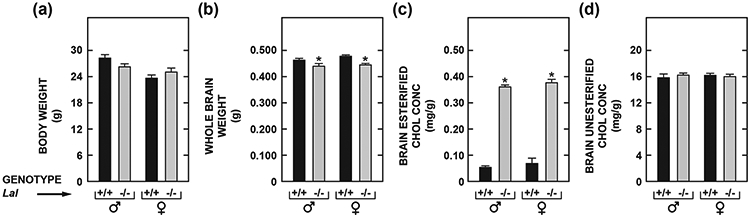
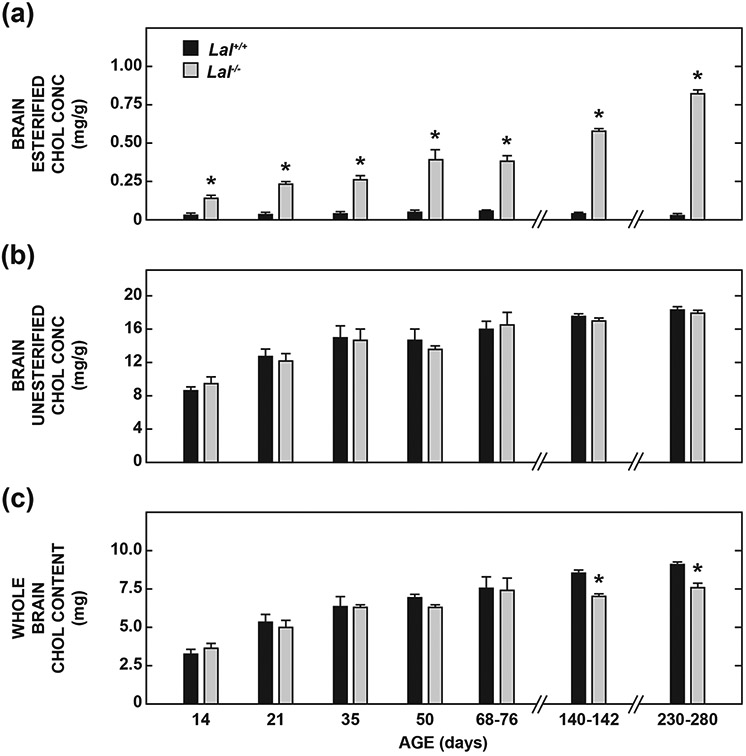
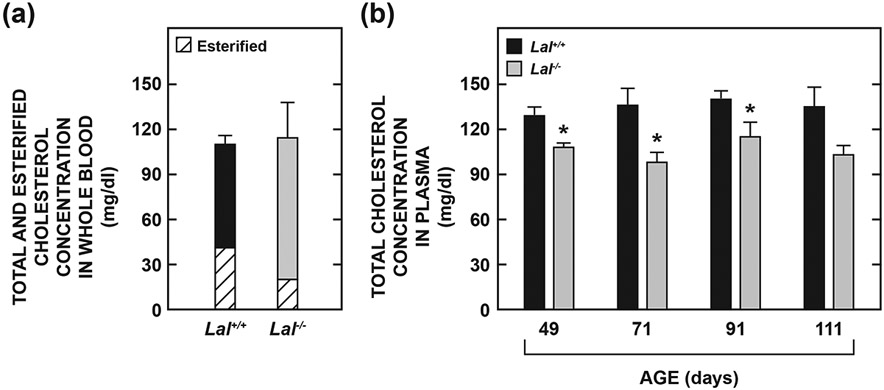
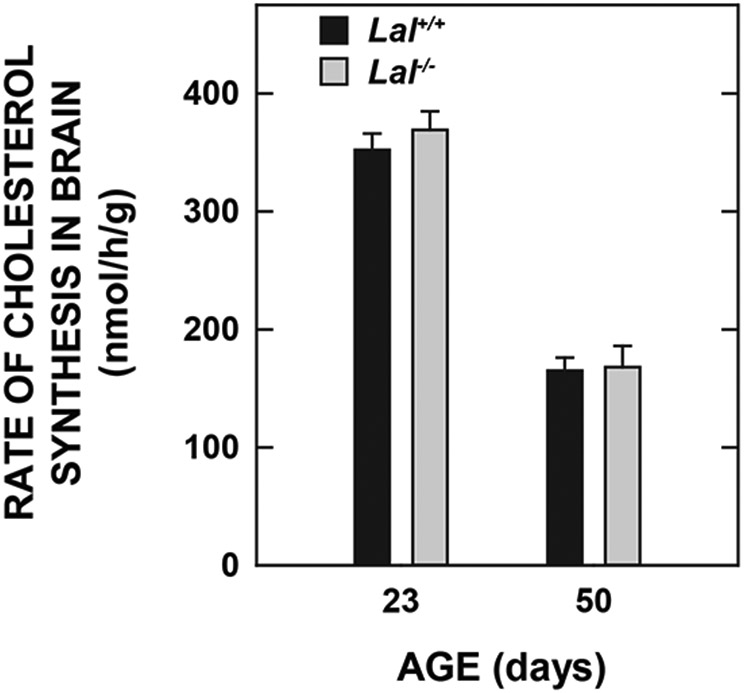
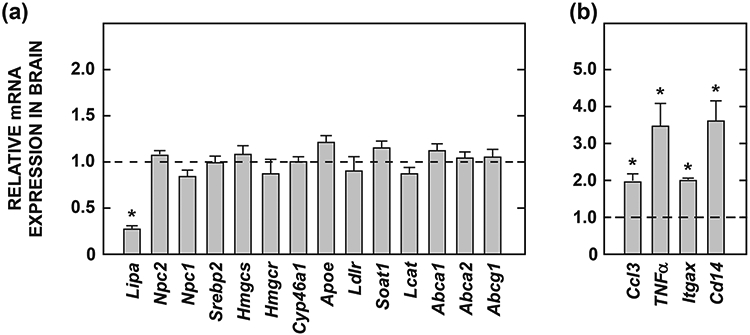
Lysosomal acid lipase (LAL), encoded by the gene LIPA, facilitates the intracellular processing of lipids by hydrolyzing cholesteryl esters and triacylglycerols present in newly internalized lipoproteins. Loss-of-function mutations in LIPA result in cholesteryl ester storage disease (CESD) or Wolman disease when mutations cause complete loss of LAL activity. Although the phenotype of a mouse CESD model has been extensively characterized, there has not been a focus on the brain at different stages of disease progression. In the current studies, whole-brain mass and the concentrations of cholesterol in both the esterified (EC) and unesterified (UC) fractions were measured in Lal-/- and matching Lal+/+ mice (FVB-N strain) at ages ranging from 14 up to 280 days after birth. Compared to Lal+/+ controls at 50, 68-76, 140-142, and 230-280 days of age, Lal-/- mice had brain weights that averaged approximately 6%, 7%, 18%, and 20% less, respectively. Brain EC levels were higher in the Lal-/- mice at every age, being elevated 27-fold at 230-280 days. Brain UC concentrations did not show a genotypic difference at any age. The elevated brain EC levels in the Lal-/- mice did not reflect EC in residual blood. An mRNA expression analysis for an array of genes involved in the synthesis, catabolism, storage, and transport of cholesterol in the brains of 141-day old mice did not detect any genotypic differences although the relative mRNA levels for several markers of inflammation were moderately elevated in the Lal-/- mice. The possible sites of EC accretion in the central nervous system are discussed.
Keywords: brain mass; cholesterol-esterifying enzymes; esterified cholesterol sequestration; lysosomal acid lipase; residual blood volume; unesterified cholesterol.
© 2021 AOCS.
Conflict of interest statement
Conflict of Interest
We declare no conflict of interest.
Figures
References
-
- Accad M, Smith SJ, Newland DL, Sanan DA, King LE Jr., Linton MF, Fazio S, Farese RV Jr. (2000) Massive xanthomatosis and altered composition of atherosclerotic lesions in hyperlipidemic mice lacking acyl CoA:cholesterol acyltransferase 1. The Journal of Clinical Investigation, 105:711–719. - PMC - PubMed
-
- Alling C, & Svennerholm L (1969) Concentration and fatty acid composition of cholesteryl esters of normal human brain. Journal of Neurochemistry, 16:751–759. - PubMed
-
- Aqul A, Liu B, Ramirez CM, Pieper AA, Estill SJ, Burns DK, Liu B, Repa JJ, Turley SD, & Dietschy JM (2011) Unesterified cholesterol accumulation in late endosomes/lysosomes causes neurodegeneration and is prevented by driving cholesterol export from this compartment. The Journal of Neuroscience, 31:9404–9413. - PMC - PubMed
-
- Aqul A, Lopez AM, Posey KS, Taylor AM, Repa JJ, Burns DK, & Turley SD (2014) Hepatic entrapment of esterified cholesterol drives continual expansion of whole body sterol pool in lysosomal acid lipase-deficient mice. American Journal of Physiology-Gastrointestinal and Liver Physiology, 307:G836–847. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
