

Cellular pathways influenced by protein arginine methylation: Implications for cancer
- PMID: 34619091
- PMCID: PMC8571027
- DOI: 10.1016/j.molcel.2021.09.011
Cellular pathways influenced by protein arginine methylation: Implications for cancer
Abstract
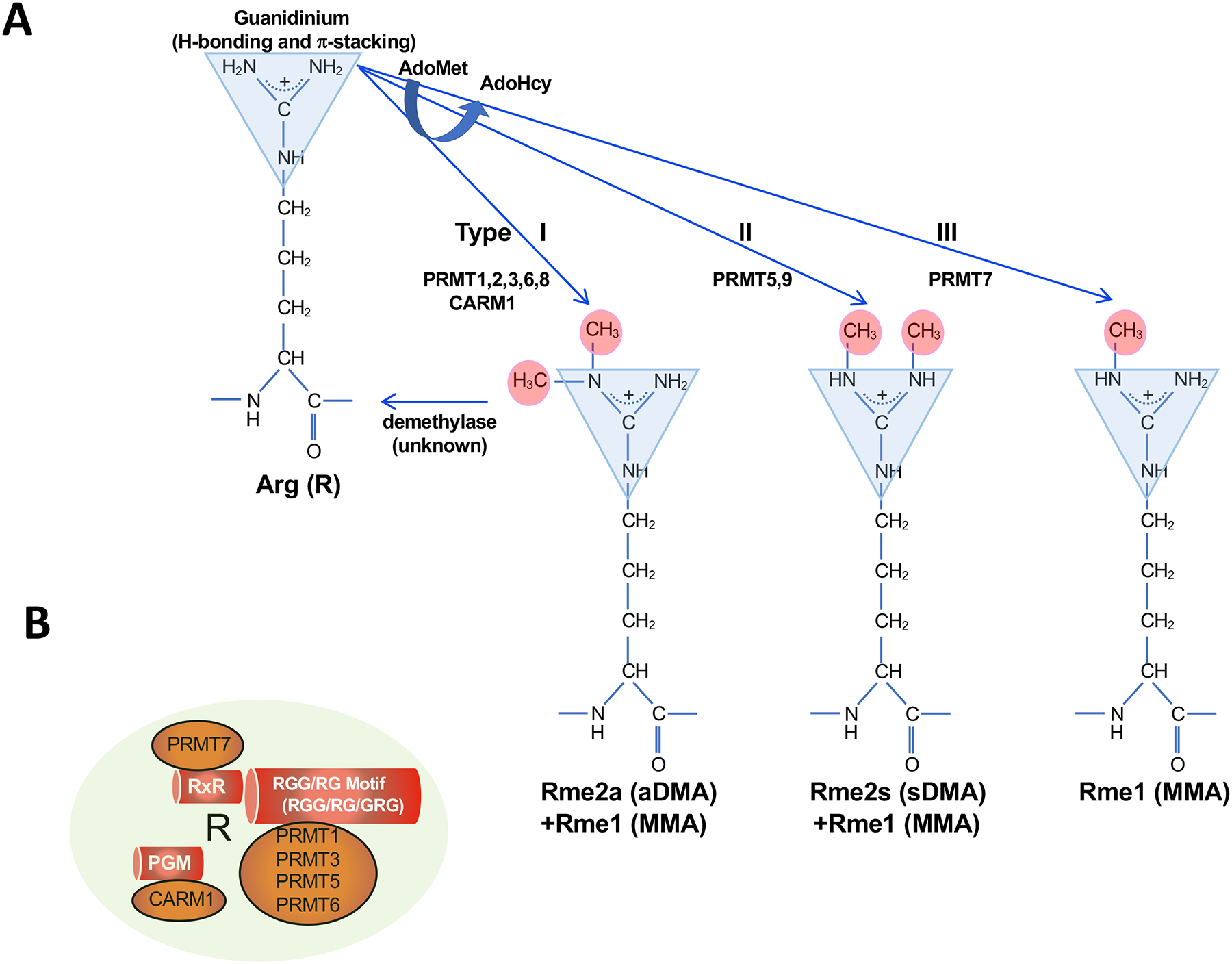
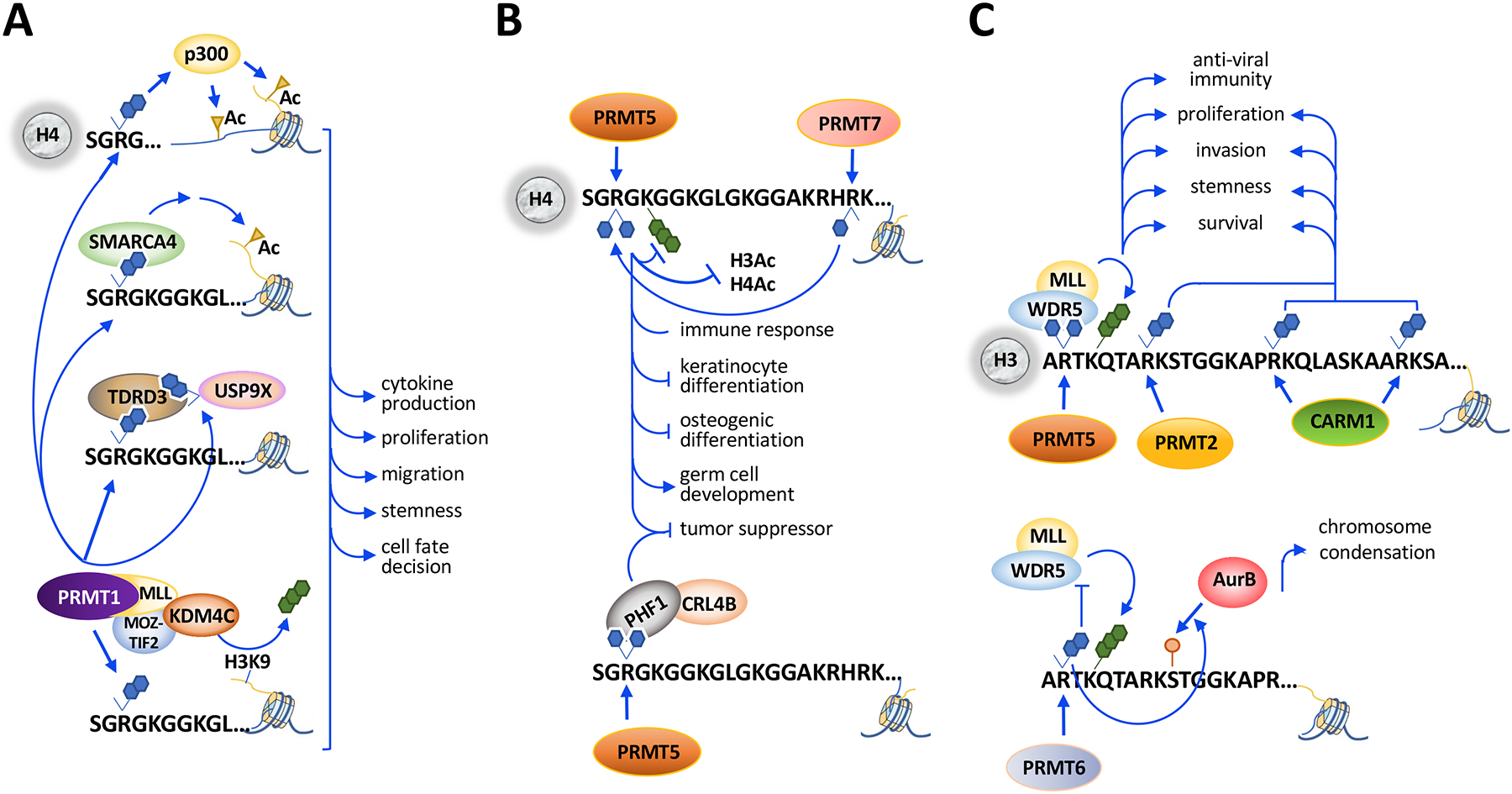
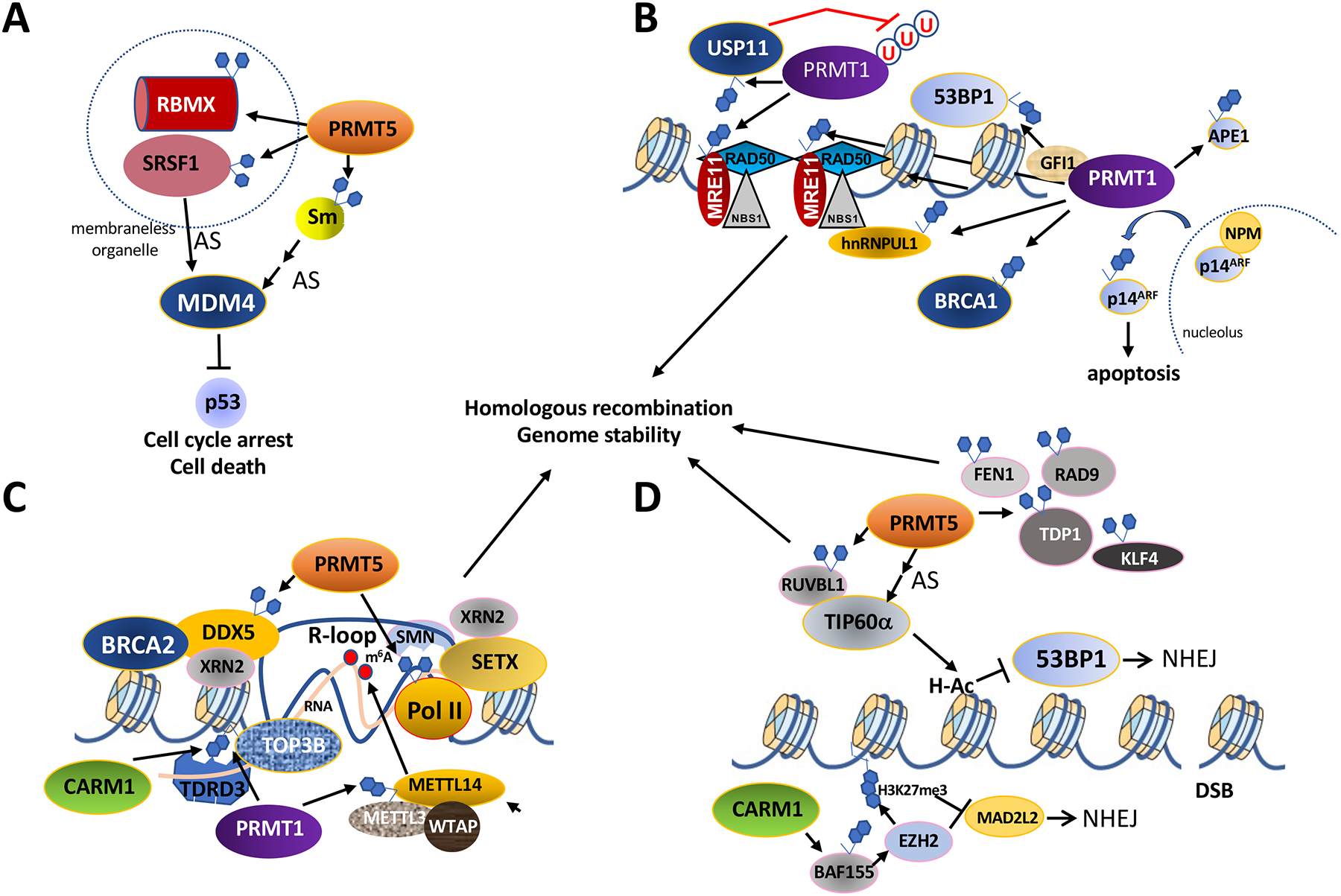
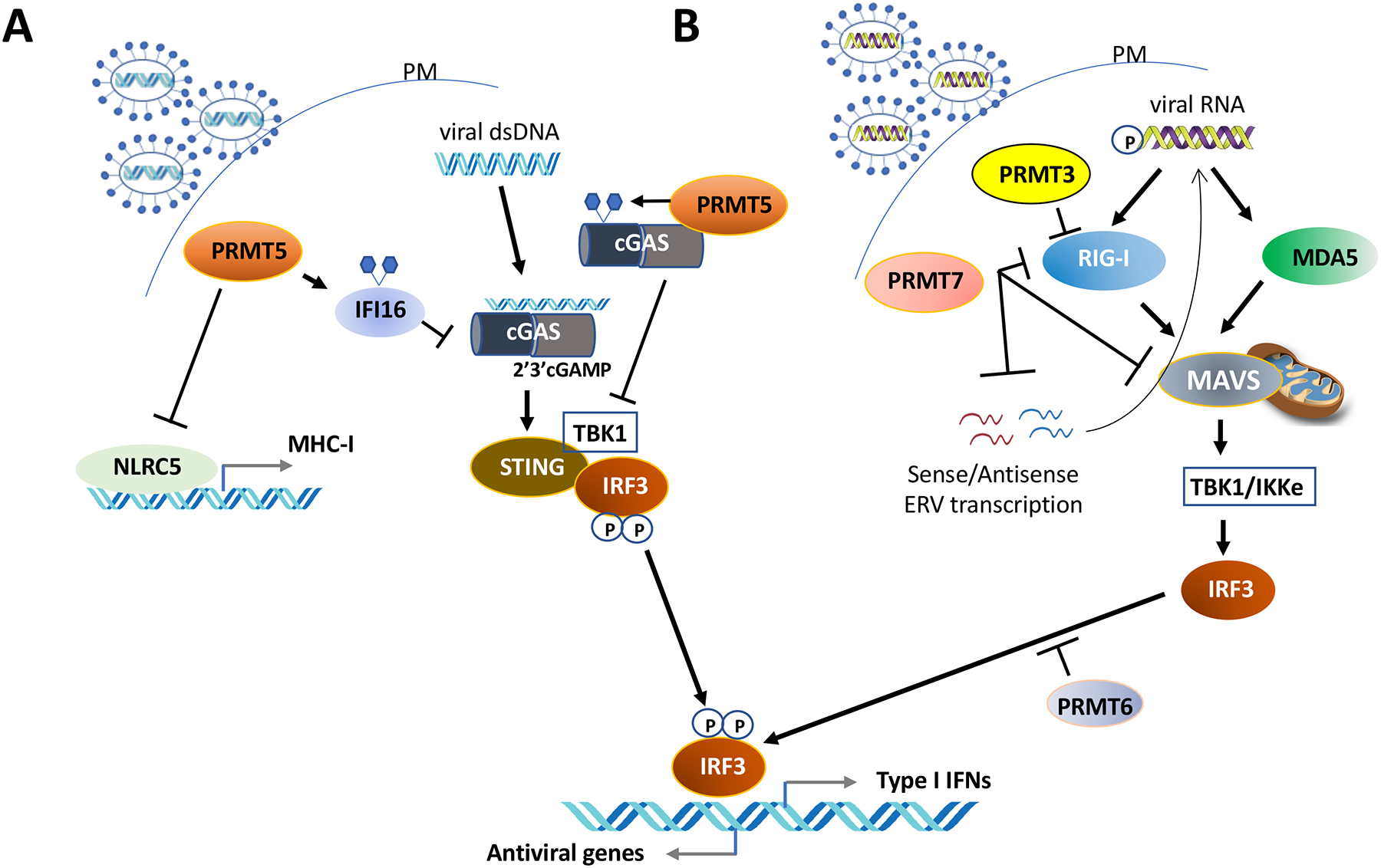
Arginine methylation is an influential post-translational modification occurring on histones, RNA binding proteins, and many other cellular proteins, affecting their function by altering their protein-protein and protein-nucleic acid interactions. Recently, a wealth of information has been gathered, implicating protein arginine methyltransferases (PRMTs), enzymes that deposit arginine methylation, in transcription, pre-mRNA splicing, DNA damage signaling, and immune signaling with major implications for cancer therapy, especially immunotherapy. This review summarizes this recent progress and the current state of PRMT inhibitors, some in clinical trials, as promising drug targets for cancer.
Keywords: DNA damage signaling; PRMTs; S-adenosylmethionine; arginine methylation; epigenetics; immunotherapy; pre-mRNA splicing; small-molecule inhibitors; transcription.
Copyright © 2021 Elsevier Inc. All rights reserved.
Figures
References
-
- Agolini E, D. M, Bellacchio E, Alesi V, Radio FC, Torella A, Musacchia F, Tartaglia M, Dallapiccola B, Nigro V, Digilio MC, Novelli A. (2018). Expanding the clinical and molecular spectrum of PRMT7 mutations: 3 additional patients and review. Clin Genet 93, 675–681. - PubMed
-
- An W, Kim J, and Roeder RG (2004). Ordered cooperative functions of PRMT1, p300, and CARM1 in transcriptional activation by p53. Cell 117, 735–748. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
