

A mutation-mediated evolutionary adaptation of Zika virus in mosquito and mammalian host
- PMID: 34620704
- PMCID: PMC8545446
- DOI: 10.1073/pnas.2113015118
A mutation-mediated evolutionary adaptation of Zika virus in mosquito and mammalian host
Abstract
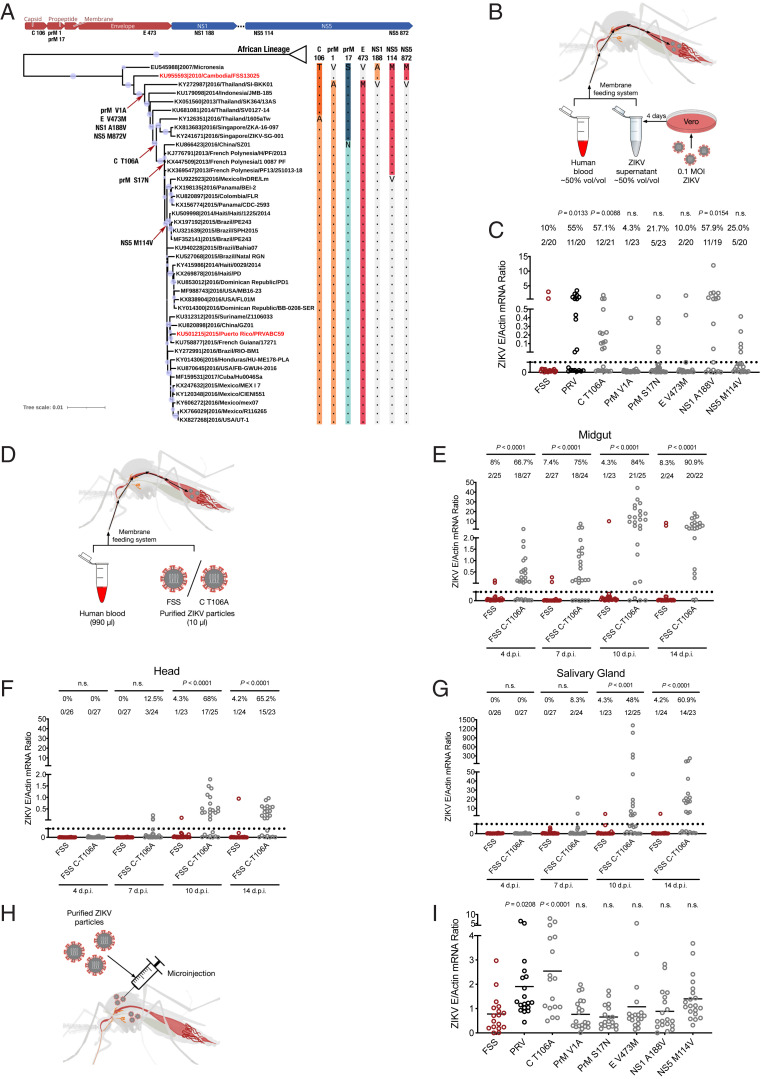
Zika virus (ZIKV) caused millions of infections during its rapid and expansive spread from Asia to the Americas from 2015 to 2017. Here, we compared the infectivity of ZIKV mutants with individual stable substitutions which emerged throughout the Asian ZIKV lineage and were responsible for the explosive outbreaks in the Americas. A threonine (T) to alanine (A) mutation at the 106th residue of the ZIKV capsid (C) protein facilitated the transmission by its mosquito vector, as well as infection in both human cells and immunodeficient mice. A mechanistic study showed that the T106A substitution rendered the C a preferred substrate for the NS2B-NS3 protease, thereby facilitating the maturation of structural proteins and the formation of infectious viral particles. Over a complete "mosquito-mouse-mosquito" cycle, the ZIKV C-T106A mutant showed a higher prevalence of mosquito infection than did the preepidemic strain, thus promoting ZIKV dissemination. Our results support the contribution of this evolutionary adaptation to the occasional widespread reemergence of ZIKV in nature.
Keywords: Zika virus; evolutionary adaptation; host; mosquito; mutation.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Musso D., Nilles E. J., Cao-Lormeau V.-M., Rapid spread of emerging Zika virus in the Pacific area. Clin. Microbiol. Infect. 20, O595–O596 (2014). - PubMed
-
- Marchette N. J., Garcia R., Rudnick A., Isolation of Zika virus from Aedes aegypti mosquitoes in Malaysia. Am. J. Trop. Med. Hyg. 18, 411–415 (1969). - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Medical
