

Reduction in gene expression noise by targeted increase in accessibility at gene loci
- PMID: 34625470
- PMCID: PMC8545487
- DOI: 10.1073/pnas.2018640118
Reduction in gene expression noise by targeted increase in accessibility at gene loci
Abstract
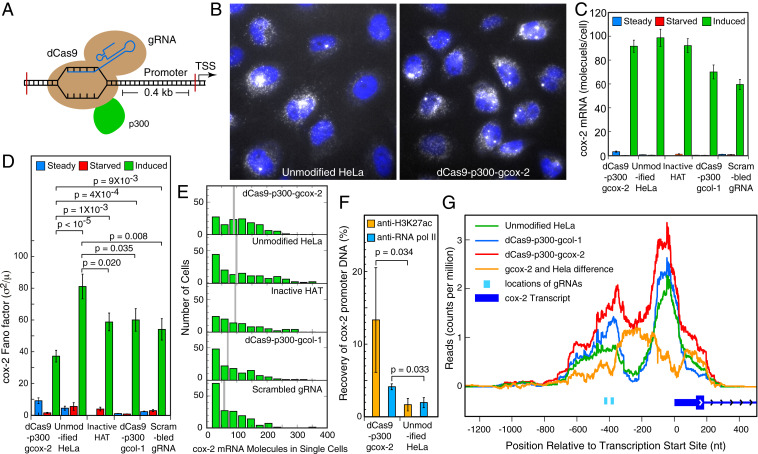
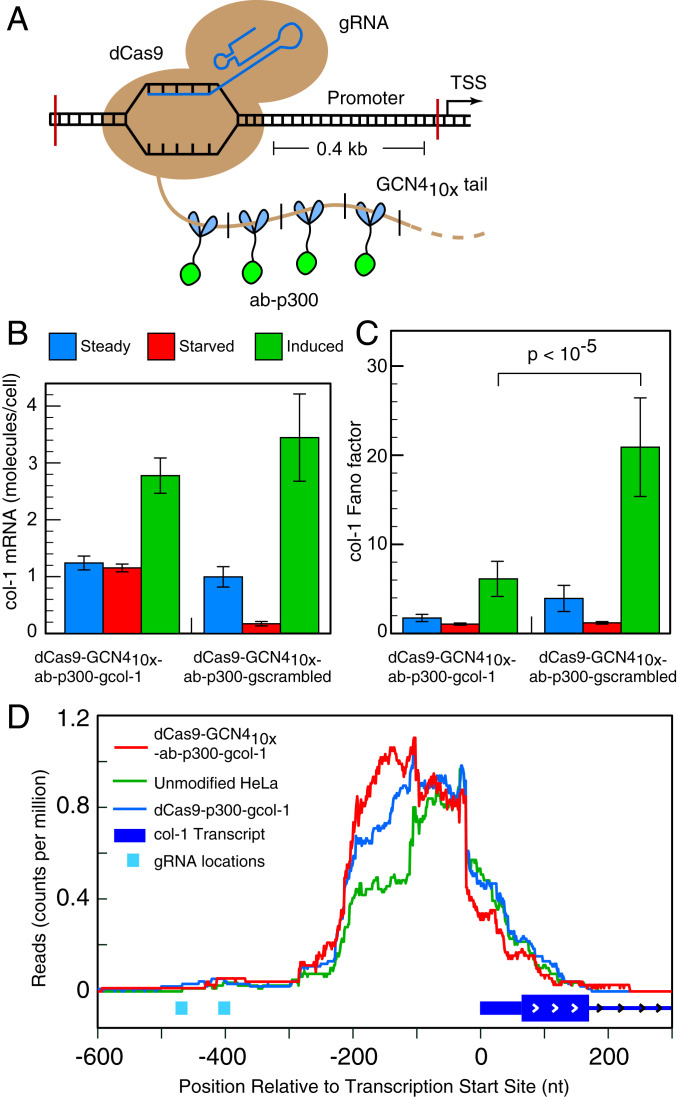
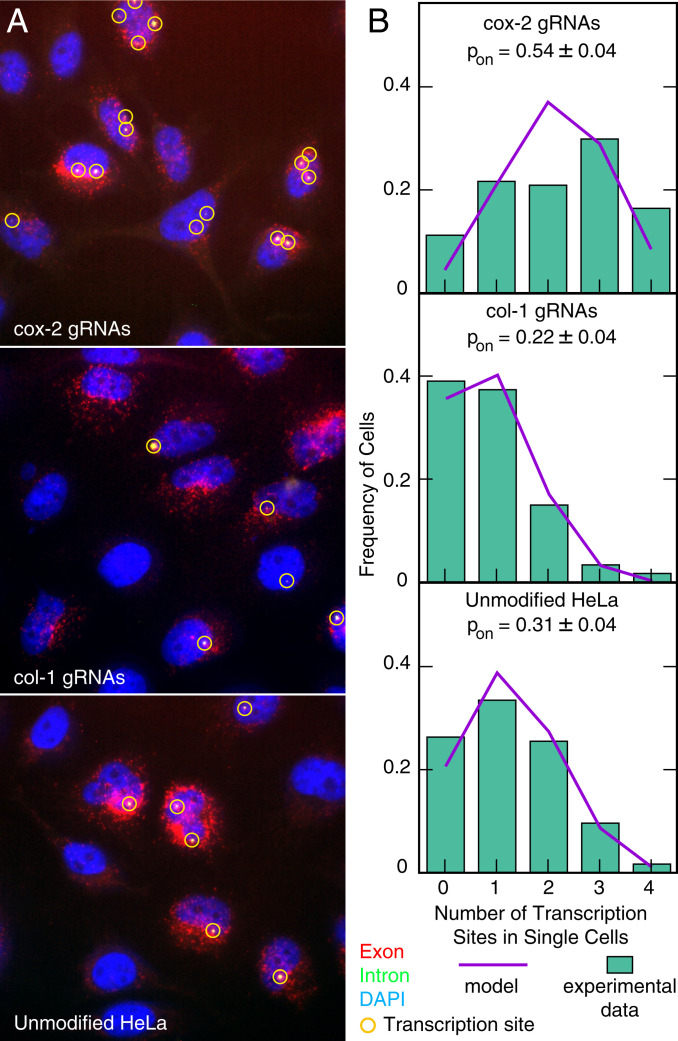
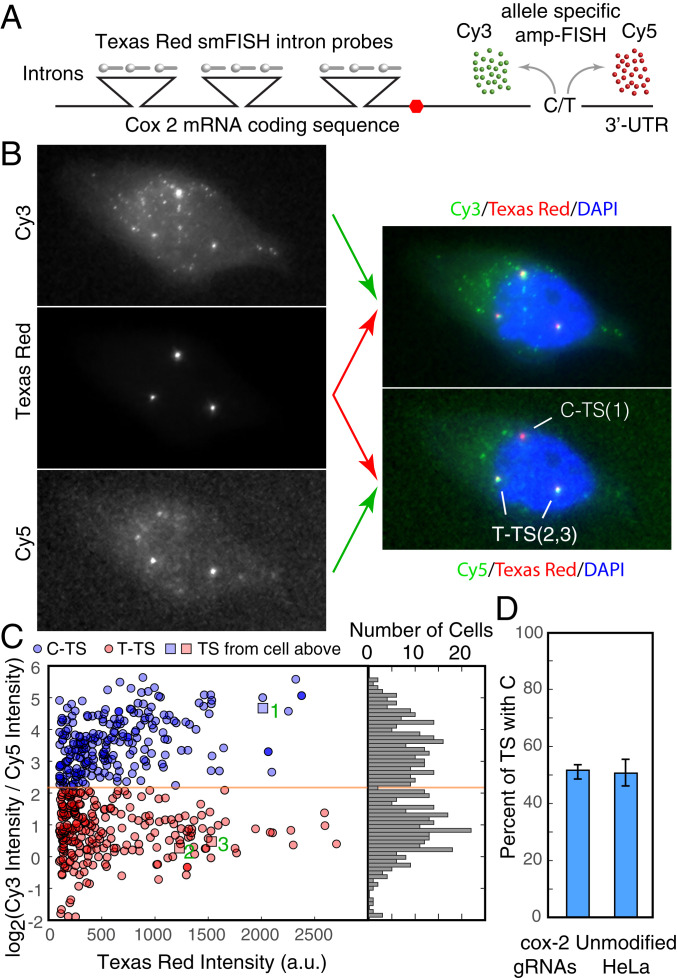
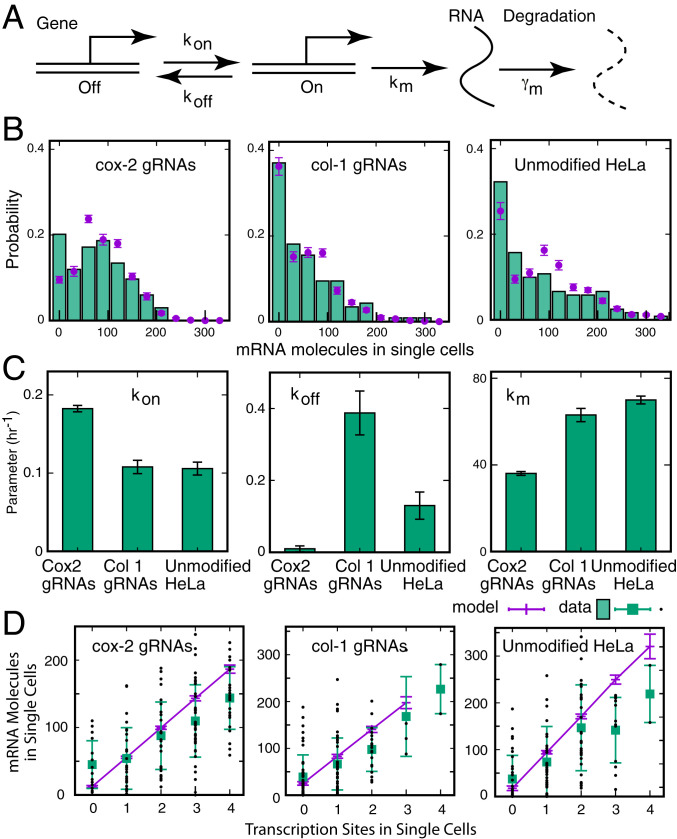
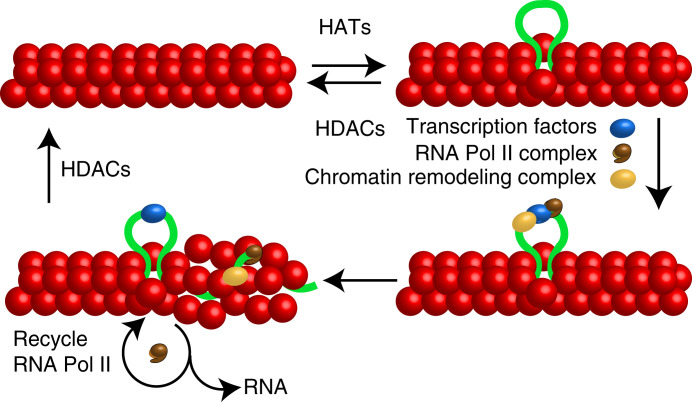
Many eukaryotic genes are expressed in randomly initiated bursts that are punctuated by periods of quiescence. Here, we show that the intermittent access of the promoters to transcription factors through relatively impervious chromatin contributes to this "noisy" transcription. We tethered a nuclease-deficient Cas9 fused to a histone acetyl transferase at the promoters of two endogenous genes in HeLa cells. An assay for transposase-accessible chromatin using sequencing showed that the activity of the histone acetyl transferase altered the chromatin architecture locally without introducing global changes in the nucleus and rendered the targeted promoters constitutively accessible. We measured the gene expression variability from the gene loci by performing single-molecule fluorescence in situ hybridization against mature messenger RNAs (mRNAs) and by imaging nascent mRNA molecules present at active gene loci in single cells. Because of the increased accessibility of the promoter to transcription factors, the transcription from two genes became less noisy, even when the average levels of expression did not change. In addition to providing evidence for chromatin accessibility as a determinant of the noise in gene expression, our study offers a mechanism for controlling gene expression noise which is otherwise unavoidable.
Keywords: single-cell heterogeneity; stochastic mRNA synthesis; transcriptional bursting.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
