

Distinct Role of Dopamine in the PFC and NAc During Exposure to Cocaine-Associated Cues
- PMID: 34626116
- PMCID: PMC8653875
- DOI: 10.1093/ijnp/pyab067
Distinct Role of Dopamine in the PFC and NAc During Exposure to Cocaine-Associated Cues
Abstract
Background: Dopamine neurotransmission plays a critical role in reward in drug abuse and drug addiction. However, the role of dopamine in the recognition of drug-associated environmental stimuli, retrieval of drug-associated memory, and drug-seeking behaviors is not fully understood.
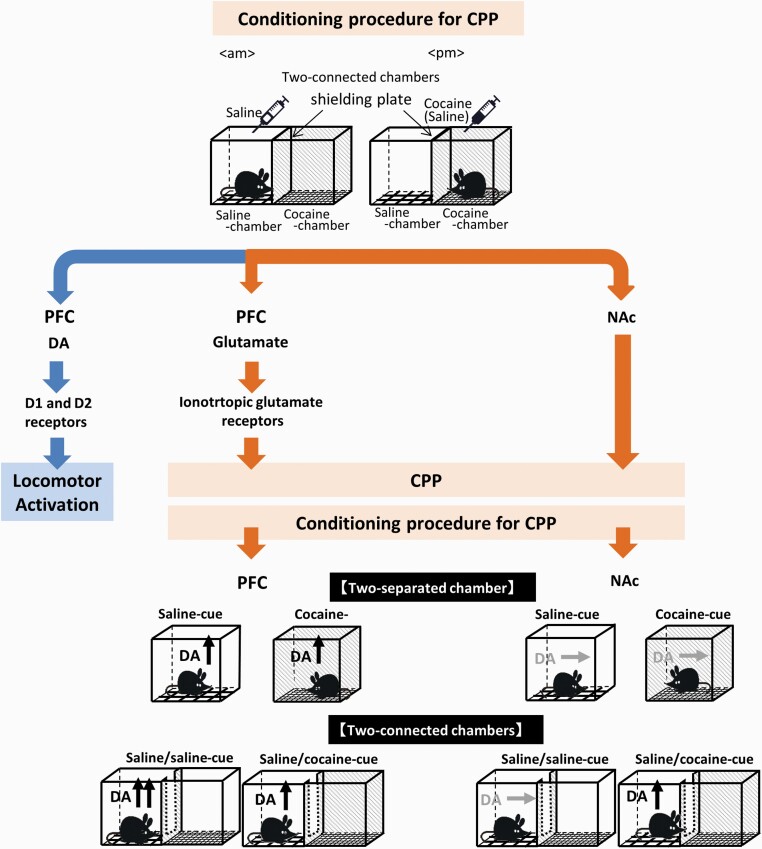
Methods: Roles of dopamine neurotransmission in the prefrontal cortex (PFC) and nucleus accumbens (NAc) in the cocaine-conditioned place preference (CPP) paradigm were evaluated using in vivo microdialysis.
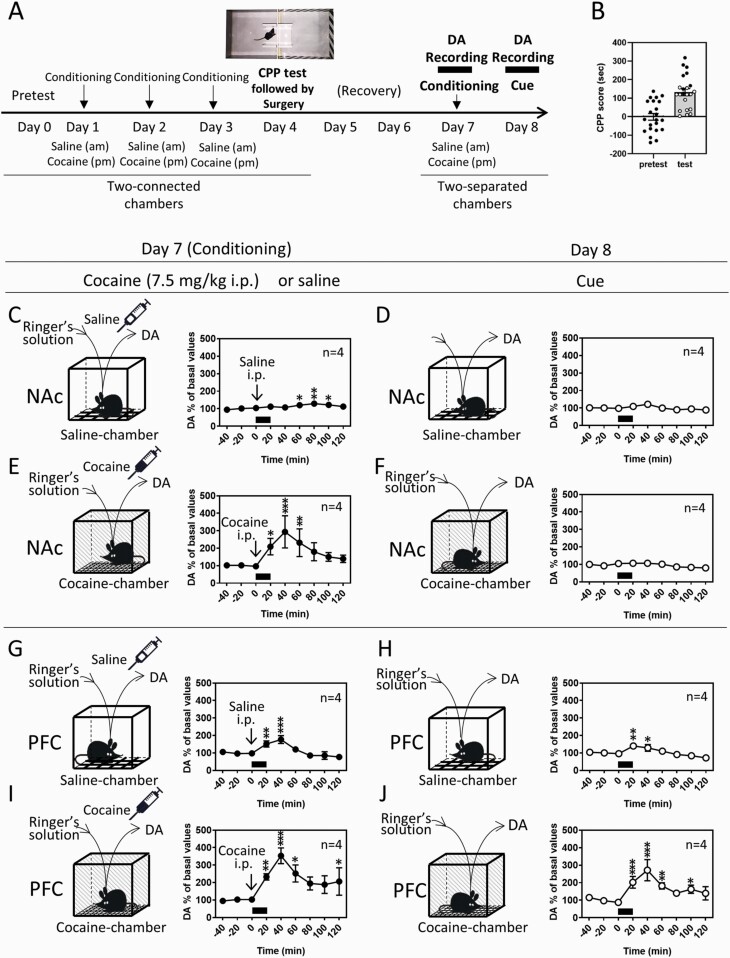
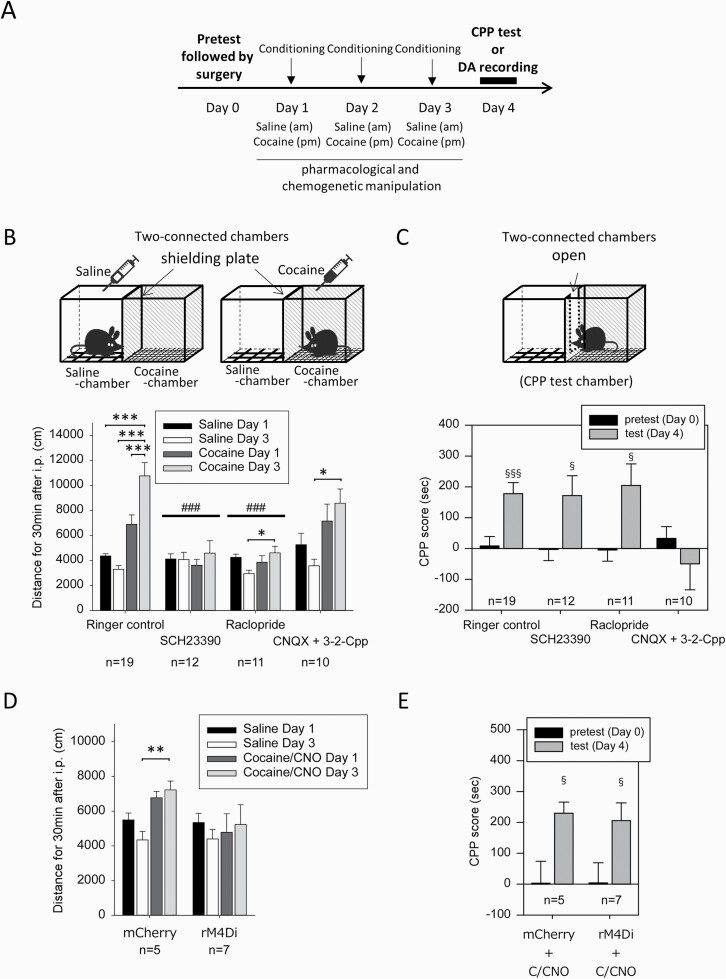
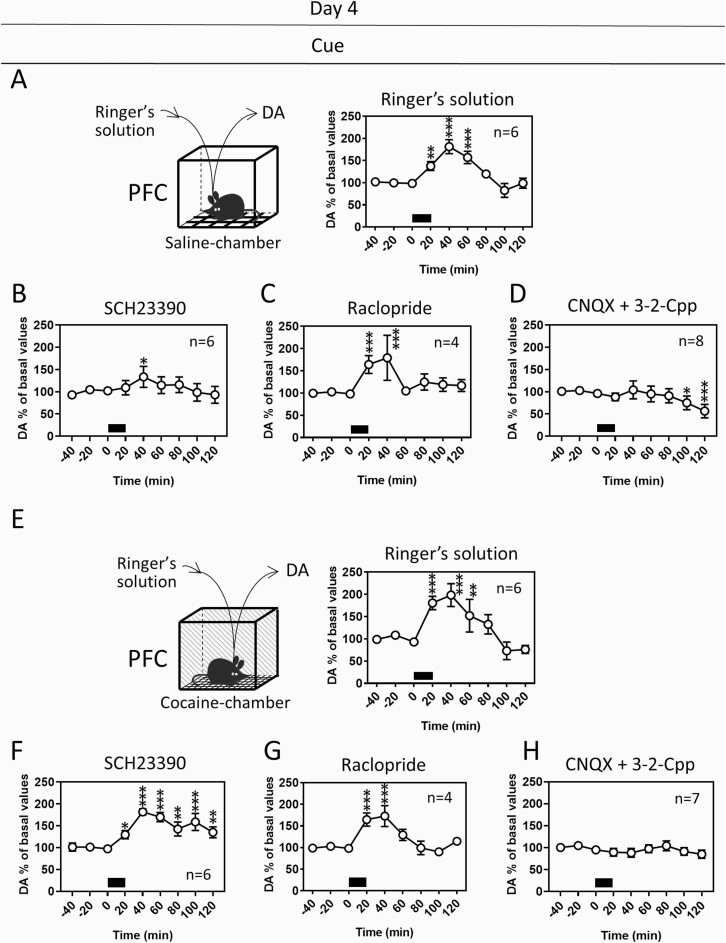
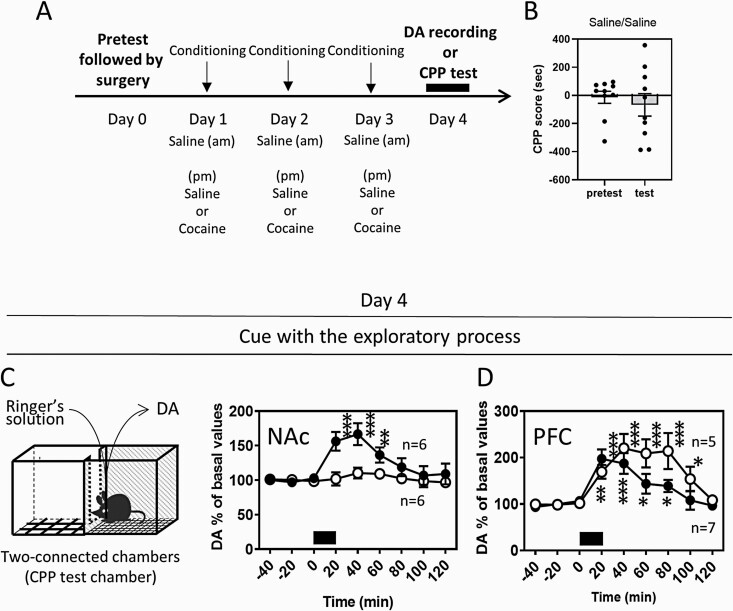
Results: In mice that had acquired cocaine CPP, dopamine levels in the PFC, but not in the NAc, increased in response to cocaine-associated cues when mice were placed in the cocaine chamber of an apparatus with 2 separated chambers. The induction of the dopamine response and the development of cocaine CPP were mediated through activation of glutamate NMDA (N-methyl-D-aspartate)/AMPA (α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid) receptor signaling in the PFC during conditioning. Activation of dopamine D1 or D2 receptor signaling in the PFC was required for cocaine-induced locomotion, but not for the induction of the dopamine response or the development of cocaine CPP. Interestingly, dopamine levels in the NAc increased in response to cocaine-associated cues when mice were placed at the center of an apparatus with 2 connected chambers, which requires motivated exploration associated with cocaine reward.
Conclusions: Dopamine neurotransmission in the PFC is activated by the exposure to the cocaine-associated cues, whereas dopamine neurotransmission in the NAc is activated in a process of motivated exploration of cues associated with cocaine reward. Furthermore, the glutamate signaling cascade in the PFC is suggested to be a potential therapeutic target to prevent the progression of drug addiction.
Keywords: Conditioned place preference; D1 receptor; D2 receptor; glutamate; microdialysis.
© The Author(s) 2021. Published by Oxford University Press on behalf of CINP.
Figures
Similar articles
-
NMDA and dopamine D1 receptors within NAc-shell regulate IEG proteins expression in reward circuit during cocaine memory reconsolidation.Neuroscience. 2016 Feb 19;315:45-69. doi: 10.1016/j.neuroscience.2015.11.063. Epub 2015 Dec 8. Neuroscience. 2016. PMID: 26674058
-
Blockade of NMDA receptors in the prefrontal cortex increases dopamine and acetylcholine release in the nucleus accumbens and motor activity.Psychopharmacology (Berl). 2008 Dec;201(3):325-38. doi: 10.1007/s00213-008-1288-3. Epub 2008 Aug 28. Psychopharmacology (Berl). 2008. PMID: 18751970
-
In vivo imaging identifies temporal signature of D1 and D2 medium spiny neurons in cocaine reward.Proc Natl Acad Sci U S A. 2016 Mar 8;113(10):2726-31. doi: 10.1073/pnas.1521238113. Epub 2016 Feb 1. Proc Natl Acad Sci U S A. 2016. PMID: 26831103 Free PMC article.
-
Prefrontal cortex-nucleus accumbens interaction: in vivo modulation by dopamine and glutamate in the prefrontal cortex.Pharmacol Biochem Behav. 2008 Aug;90(2):226-35. doi: 10.1016/j.pbb.2008.04.011. Epub 2008 Apr 23. Pharmacol Biochem Behav. 2008. PMID: 18508116 Review.
-
Role of nucleus accumbens glutamatergic plasticity in drug addiction.Neuropsychiatr Dis Treat. 2013;9:1499-512. doi: 10.2147/NDT.S45963. Epub 2013 Sep 30. Neuropsychiatr Dis Treat. 2013. PMID: 24109187 Free PMC article. Review.
Cited by
-
Corticostriatal ensemble dynamics across heroin self-administration to reinstatement.bioRxiv [Preprint]. 2024 Jun 27:2024.06.21.599790. doi: 10.1101/2024.06.21.599790. bioRxiv. 2024. PMID: 38979314 Free PMC article. Preprint.
-
Cocaine: An Updated Overview on Chemistry, Detection, Biokinetics, and Pharmacotoxicological Aspects including Abuse Pattern.Toxins (Basel). 2022 Apr 13;14(4):278. doi: 10.3390/toxins14040278. Toxins (Basel). 2022. PMID: 35448887 Free PMC article. Review.
-
Neurobiological mechanisms and related clinical treatment of addiction: a review.Psychoradiology. 2022 Dec 16;2(4):180-189. doi: 10.1093/psyrad/kkac021. eCollection 2022 Dec. Psychoradiology. 2022. PMID: 38665277 Free PMC article. Review.
-
Immediate glucose signaling transmitted via the vagus nerve in gut-brain neural communication.iScience. 2025 May 5;28(5):112439. doi: 10.1016/j.isci.2025.112439. eCollection 2025 May 16. iScience. 2025. PMID: 40491956 Free PMC article.
-
Enhanced Beta2-band Oscillations Denote Auditory Hallucination in Schizophrenia Patients and a Monkey Model of Psychosis.Neurosci Bull. 2024 Mar;40(3):325-338. doi: 10.1007/s12264-023-01100-2. Epub 2023 Aug 24. Neurosci Bull. 2024. PMID: 37612582 Free PMC article.
References
-
- Airavaara M, Planken A, Gäddnäs H, Piepponen TP, Saarma M, Ahtee L (2004) Increased extracellular dopamine concentrations and FosB/DeltaFosB expression in striatal brain areas of heterozygous GDNF knockout mice. Eur J Neurosci 20:2336–2344. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
