

Breast cancer growth and proliferation is suppressed by the mitochondrial targeted furazano[3,4-b]pyrazine BAM15
- PMID: 34627389
- PMCID: PMC8502397
- DOI: 10.1186/s40170-021-00274-5
Breast cancer growth and proliferation is suppressed by the mitochondrial targeted furazano[3,4-b]pyrazine BAM15
Abstract
Background: Enhanced metabolic plasticity and diversification of energy production is a hallmark of highly proliferative breast cancers. This contributes to poor pharmacotherapy efficacy, recurrence, and metastases. We have previously identified a mitochondrial-targeted furazano[3,4-b]pyrazine named BAM15 that selectively reduces bioenergetic coupling efficiency and is orally available. Here, we evaluated the antineoplastic properties of uncoupling oxidative phosphorylation from ATP production in breast cancer using BAM15.
Methods: The anticancer effects of BAM15 were evaluated in human triple-negative MDA-MB-231 and murine luminal B, ERα-negative EO771 cells as well as in an orthotopic allograft model of highly proliferative mammary cancer in mice fed a standard or high fat diet (HFD). Untargeted transcriptomic profiling of MDA-MB-231 cells was conducted after 16-h exposure to BAM15. Additionally, oxidative phosphorylation and electron transfer capacity was determined in permeabilized cells and excised tumor homogenates after treatment with BAM15.
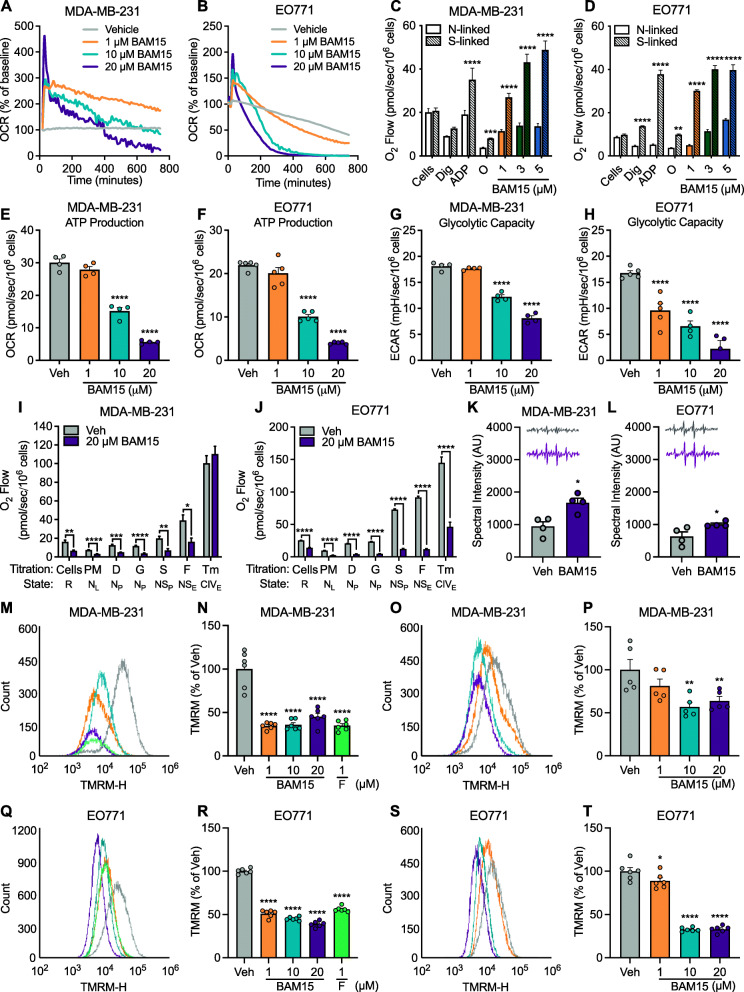
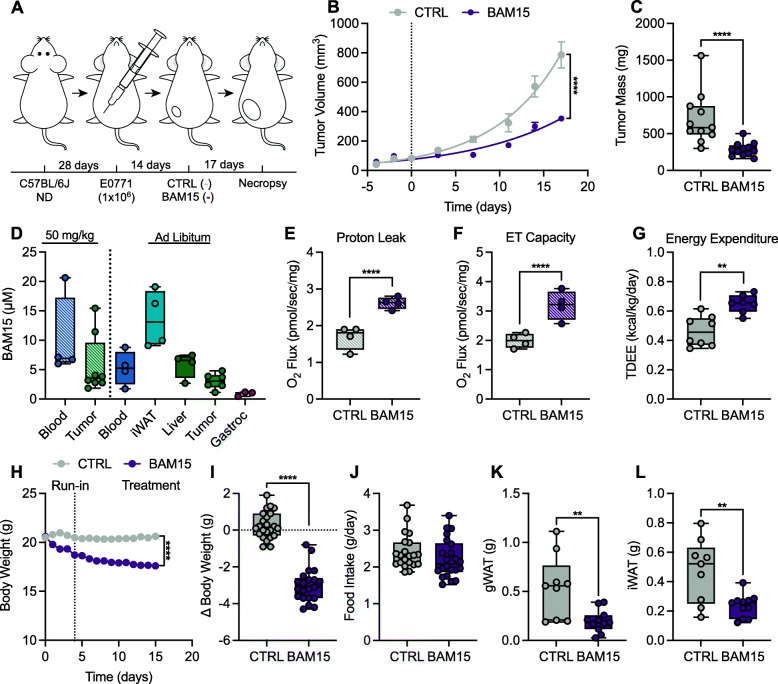
Results: BAM15 increased proton leak and over time, diminished cell proliferation, migration, and ATP production in both MDA-MB-231 and EO771 cells. Additionally, BAM15 decreased mitochondrial membrane potential, while inducing apoptosis and reactive oxygen species accumulation in MDA-MB-231 and EO771 cells. Untargeted transcriptomic profiling of MDA-MB-231 cells further revealed inhibition of signatures associated with cell survival and energy production by BAM15. In lean mice, BAM15 lowered body weight independent of food intake and slowed tumor progression compared to vehicle-treated controls. In HFD mice, BAM15 reduced tumor growth relative to vehicle and calorie-restricted weight-matched controls mediated in part by impaired cell proliferation, mitochondrial respiratory function, and ATP production. LC-MS/MS profiling of plasma and tissues from BAM15-treated animals revealed distribution of BAM15 in adipose, liver, and tumor tissue with low abundance in skeletal muscle.
Conclusions: Collectively, these data indicate that mitochondrial uncoupling may be an effective strategy to limit proliferation of aggressive forms of breast cancer. More broadly, these findings highlight the metabolic vulnerabilities of highly proliferative breast cancers which may be leveraged in overcoming poor responsiveness to existing therapies.
Keywords: BAM15; Breast cancer; Cell proliferation; Mitochondrial function; Tumor metabolism.
© 2021. The Author(s).
Conflict of interest statement
K.L.H. is an equity holder in Life Biosciences, a company that has commercial interest in mitochondrial uncouplers as therapeutics. All other authors declare no competing financial interests.
Figures




References
-
- Brown M, Tsodikov A, Bauer KR, Parise CA, Caggiano V. The role of human epidermal growth factor receptor 2 in the survival of women with estrogen and progesterone receptor-negative, invasive breast cancer: the California Cancer Registry, 1999-2004. Cancer. 2008;112(4):737–747. doi: 10.1002/cncr.23243. - DOI - PubMed
-
- Carey LA, Perou CM, Livasy CA, Dressler LG, Cowan D, Conway K, Karaca G, Troester MA, Tse CK, Edmiston S, Deming SL, Geradts J, Cheang MCU, Nielsen TO, Moorman PG, Earp HS, Millikan RC. Race, breast cancer subtypes, and survival in the Carolina Breast Cancer Study. Jama. 2006;295(21):2492–2502. doi: 10.1001/jama.295.21.2492. - DOI - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
