

Dynamic large-scale connectivity of intrinsic cortical oscillations supports adaptive listening in challenging conditions
- PMID: 34634031
- PMCID: PMC8530332
- DOI: 10.1371/journal.pbio.3001410
Dynamic large-scale connectivity of intrinsic cortical oscillations supports adaptive listening in challenging conditions
Abstract
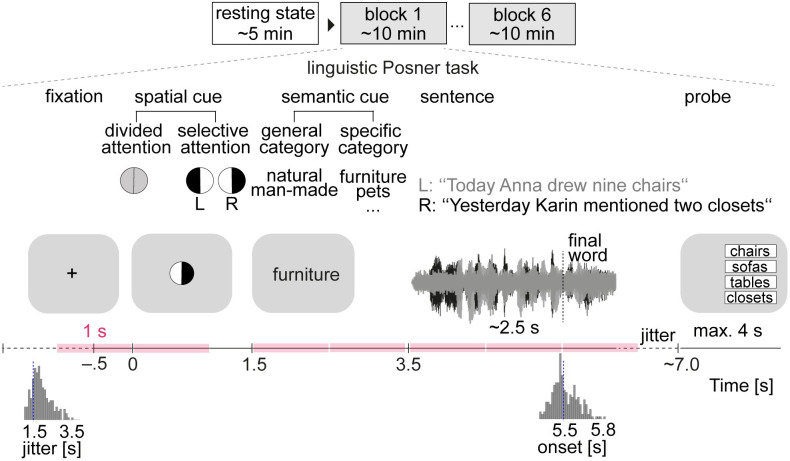
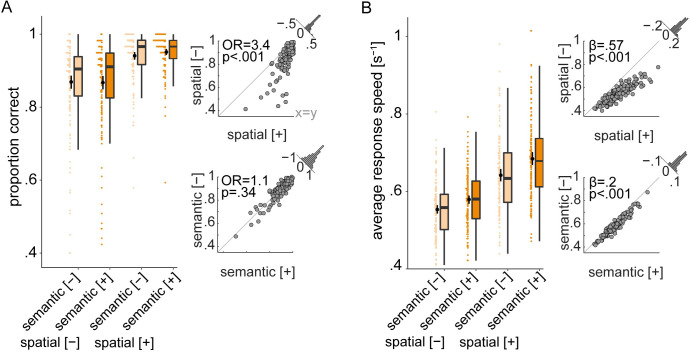
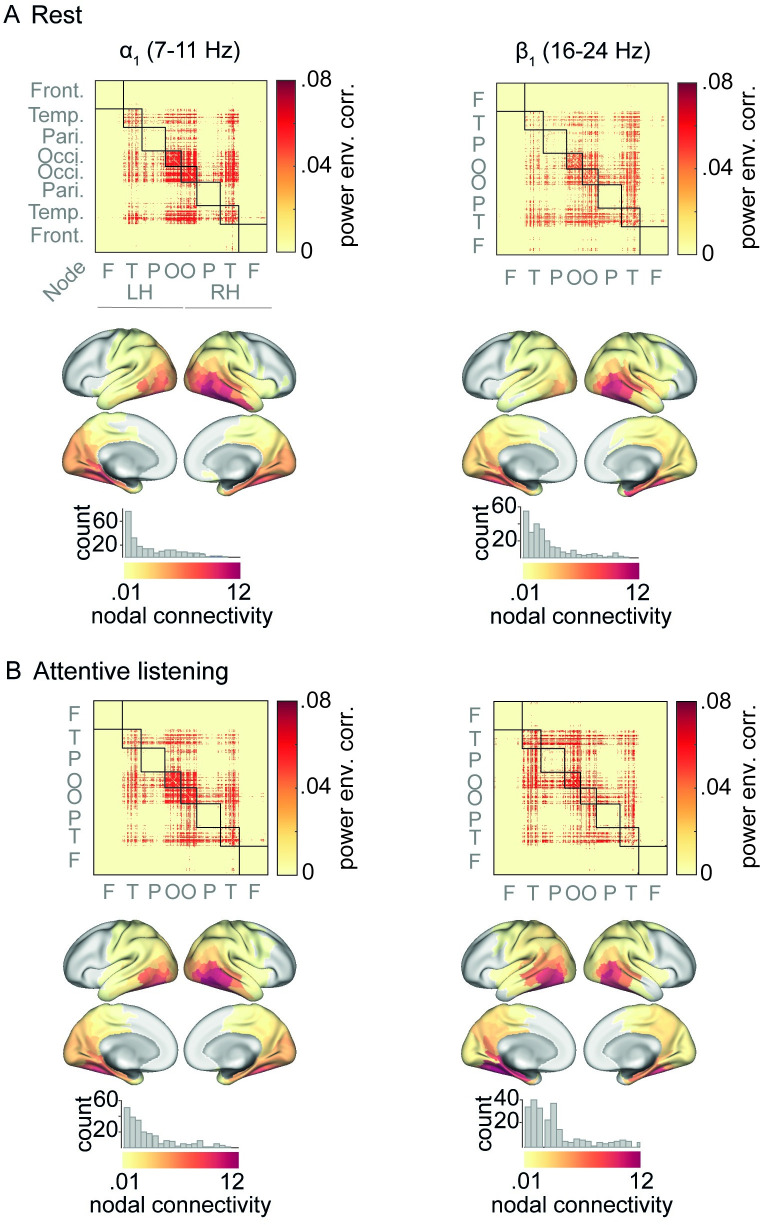
In multi-talker situations, individuals adapt behaviorally to this listening challenge mostly with ease, but how do brain neural networks shape this adaptation? We here establish a long-sought link between large-scale neural communications in electrophysiology and behavioral success in the control of attention in difficult listening situations. In an age-varying sample of N = 154 individuals, we find that connectivity between intrinsic neural oscillations extracted from source-reconstructed electroencephalography is regulated according to the listener's goal during a challenging dual-talker task. These dynamics occur as spatially organized modulations in power-envelope correlations of alpha and low-beta neural oscillations during approximately 2-s intervals most critical for listening behavior relative to resting-state baseline. First, left frontoparietal low-beta connectivity (16 to 24 Hz) increased during anticipation and processing of a spatial-attention cue before speech presentation. Second, posterior alpha connectivity (7 to 11 Hz) decreased during comprehension of competing speech, particularly around target-word presentation. Connectivity dynamics of these networks were predictive of individual differences in the speed and accuracy of target-word identification, respectively, but proved unconfounded by changes in neural oscillatory activity strength. Successful adaptation to a listening challenge thus latches onto two distinct yet complementary neural systems: a beta-tuned frontoparietal network enabling the flexible adaptation to attentive listening state and an alpha-tuned posterior network supporting attention to speech.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures



References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
