

SARS-CoV-2 nucleocapsid protein forms condensates with viral genomic RNA
- PMID: 34634033
- PMCID: PMC8553124
- DOI: 10.1371/journal.pbio.3001425
SARS-CoV-2 nucleocapsid protein forms condensates with viral genomic RNA
Abstract
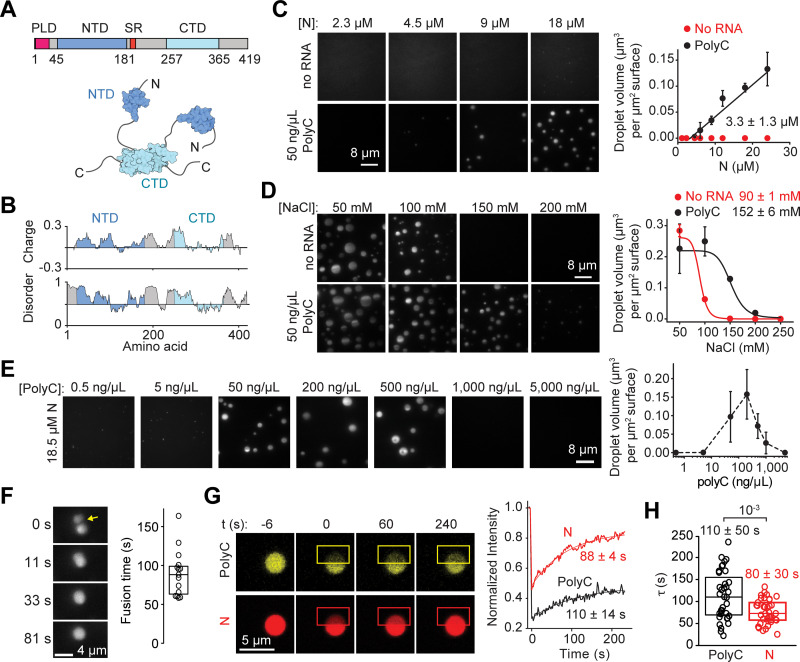
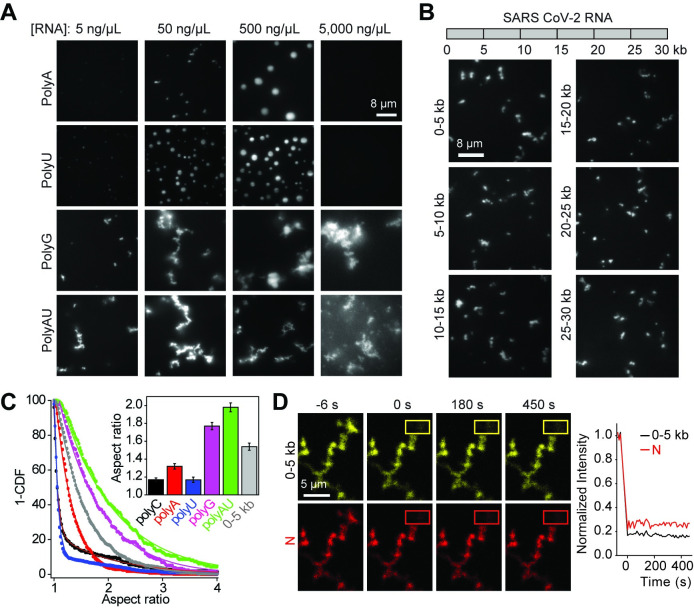
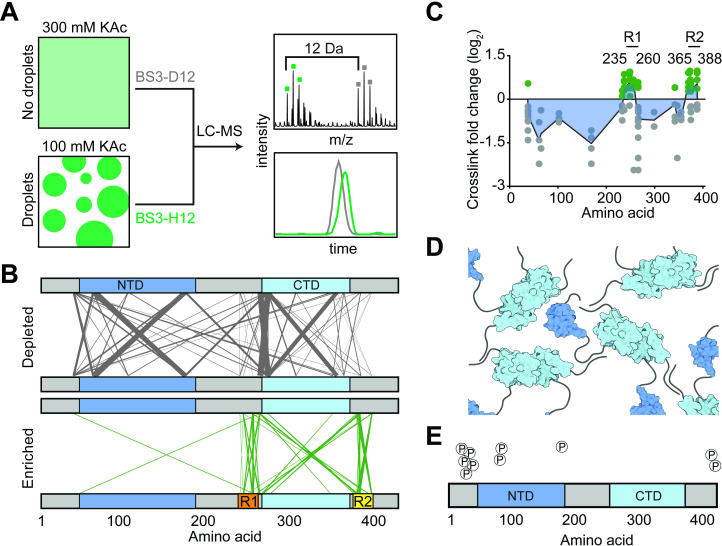
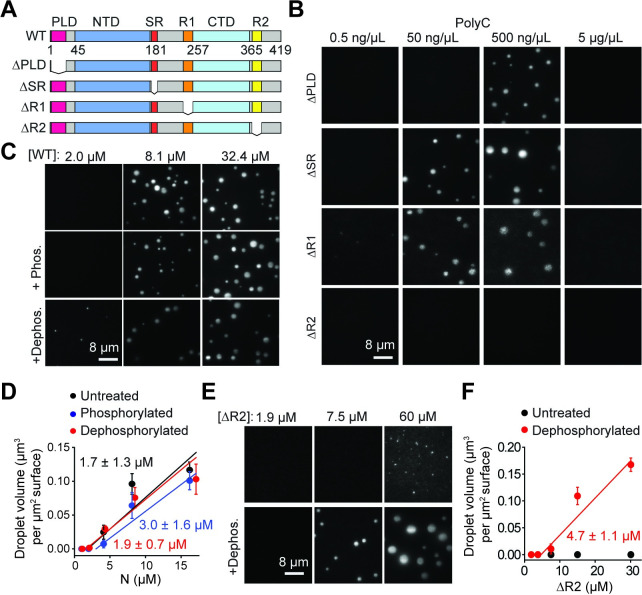
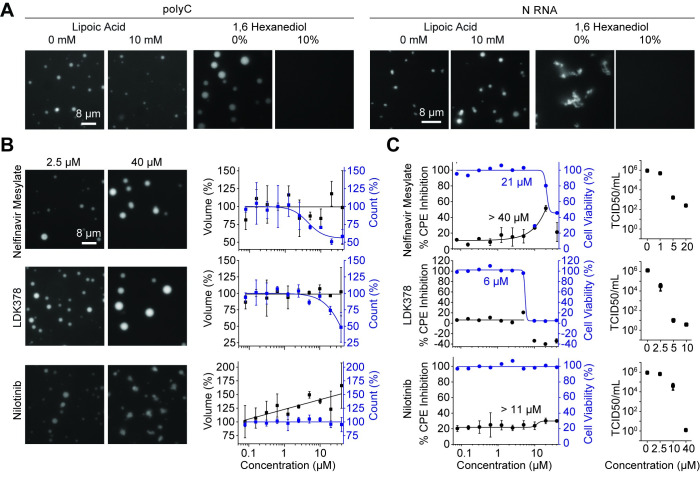
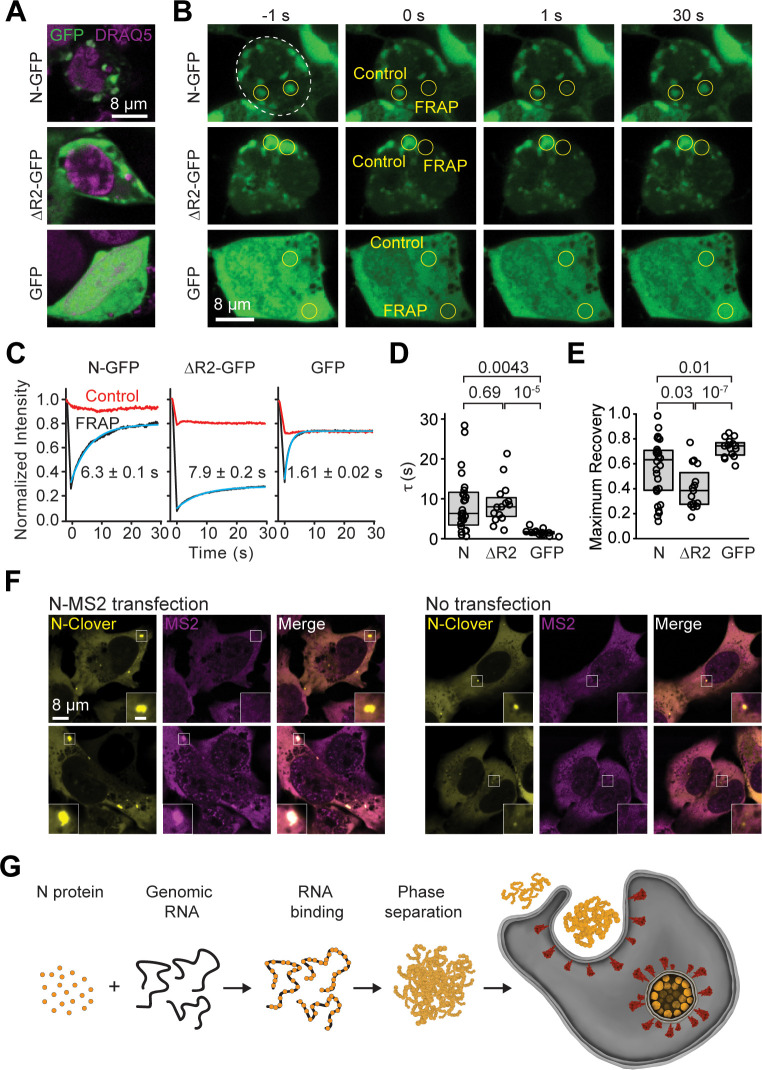
The Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) infection causes Coronavirus Disease 2019 (COVID-19), a pandemic that seriously threatens global health. SARS-CoV-2 propagates by packaging its RNA genome into membrane enclosures in host cells. The packaging of the viral genome into the nascent virion is mediated by the nucleocapsid (N) protein, but the underlying mechanism remains unclear. Here, we show that the N protein forms biomolecular condensates with viral genomic RNA both in vitro and in mammalian cells. While the N protein forms spherical assemblies with homopolymeric RNA substrates that do not form base pairing interactions, it forms asymmetric condensates with viral RNA strands. Cross-linking mass spectrometry (CLMS) identified a region that drives interactions between N proteins in condensates, and deletion of this region disrupts phase separation. We also identified small molecules that alter the size and shape of N protein condensates and inhibit the proliferation of SARS-CoV-2 in infected cells. These results suggest that the N protein may utilize biomolecular condensation to package the SARS-CoV-2 RNA genome into a viral particle.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Update of
-
SARS-CoV-2 nucleocapsid protein forms condensates with viral genomic RNA.bioRxiv [Preprint]. 2021 Mar 29:2020.09.14.295824. doi: 10.1101/2020.09.14.295824. bioRxiv. 2021. Update in: PLoS Biol. 2021 Oct 11;19(10):e3001425. doi: 10.1371/journal.pbio.3001425. PMID: 32995779 Free PMC article. Updated. Preprint.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
