

The Cyclin Cln1 Controls Polyploid Titan Cell Formation following a Stress-Induced G2 Arrest in Cryptococcus
- PMID: 34634930
- PMCID: PMC8510536
- DOI: 10.1128/mBio.02509-21
The Cyclin Cln1 Controls Polyploid Titan Cell Formation following a Stress-Induced G2 Arrest in Cryptococcus
Abstract
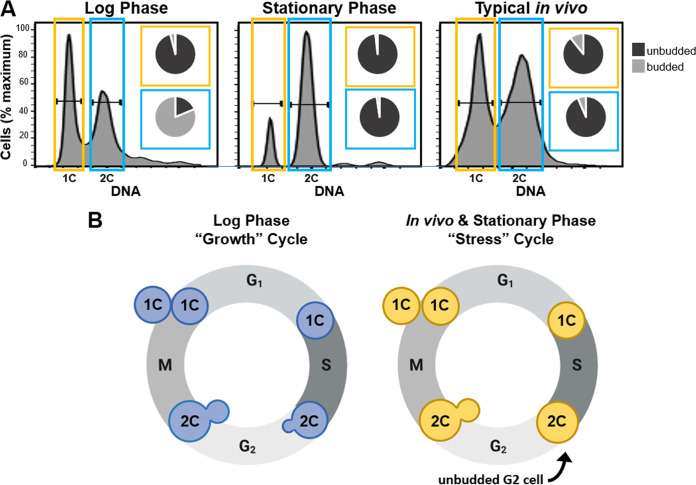
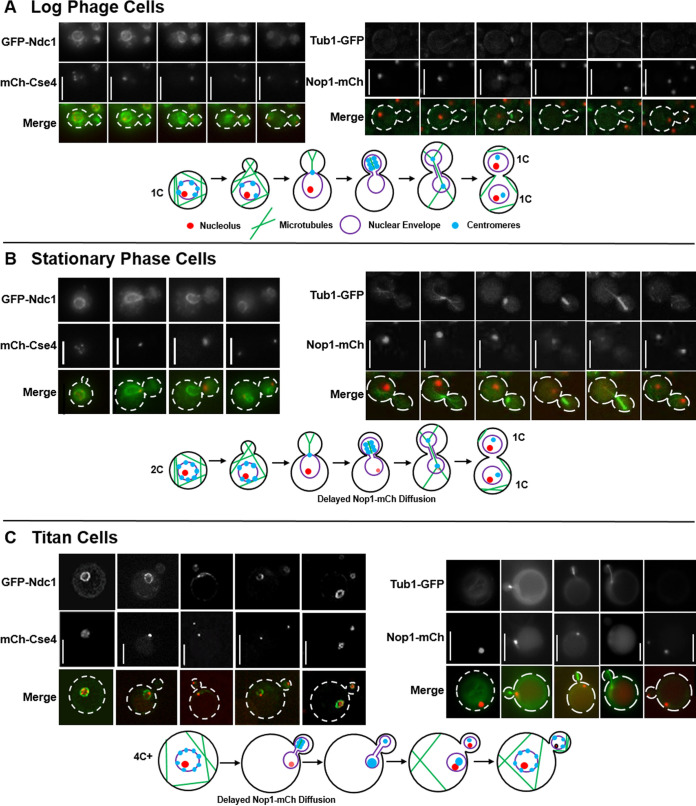
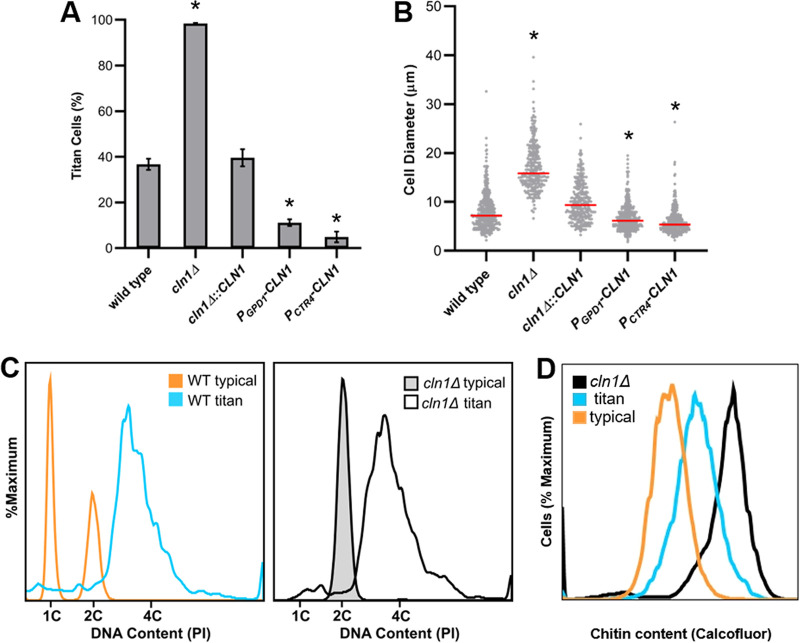
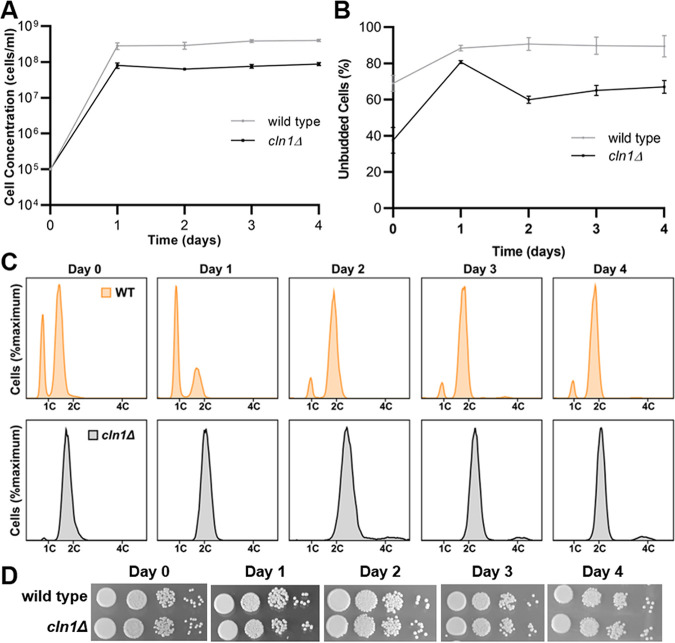
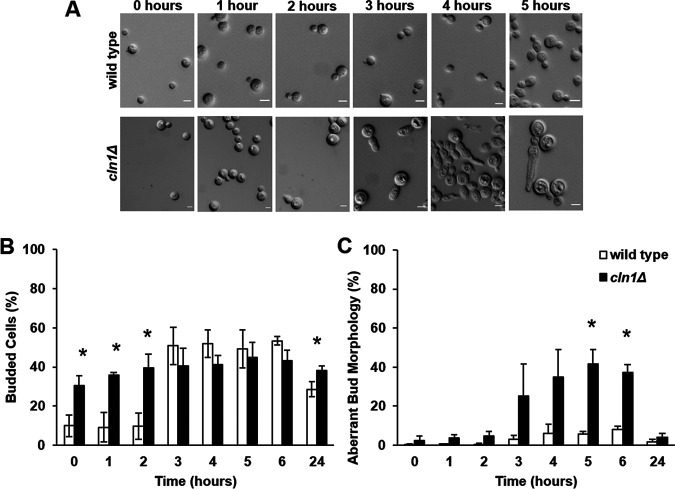
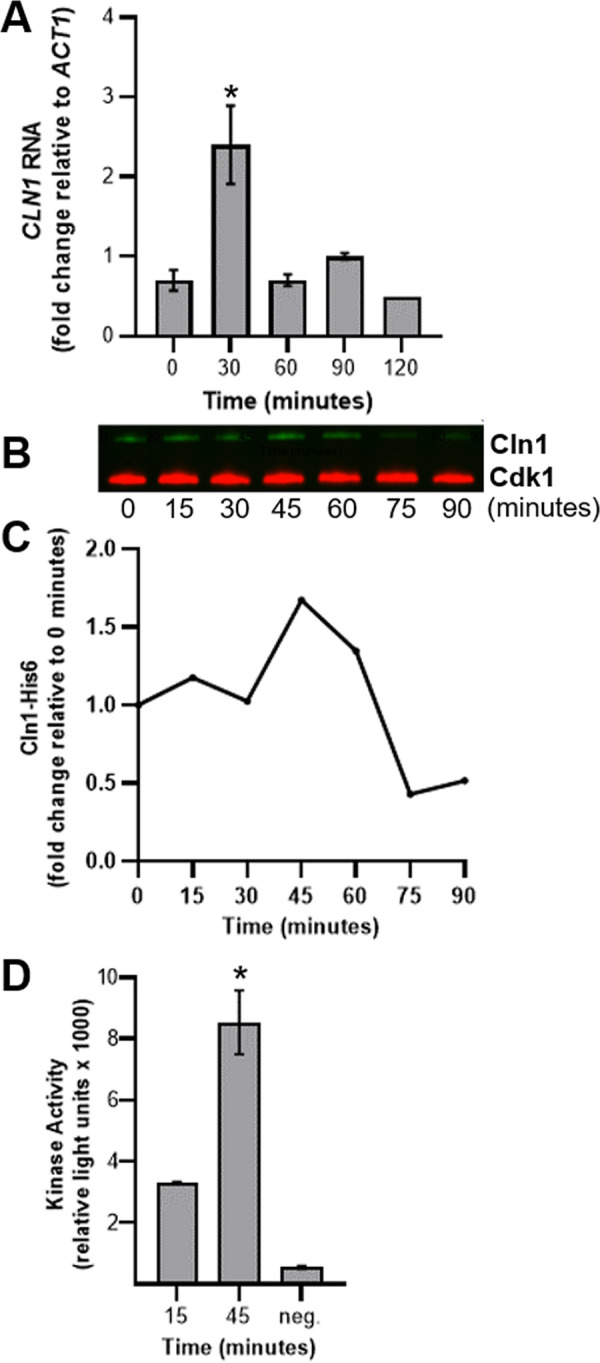
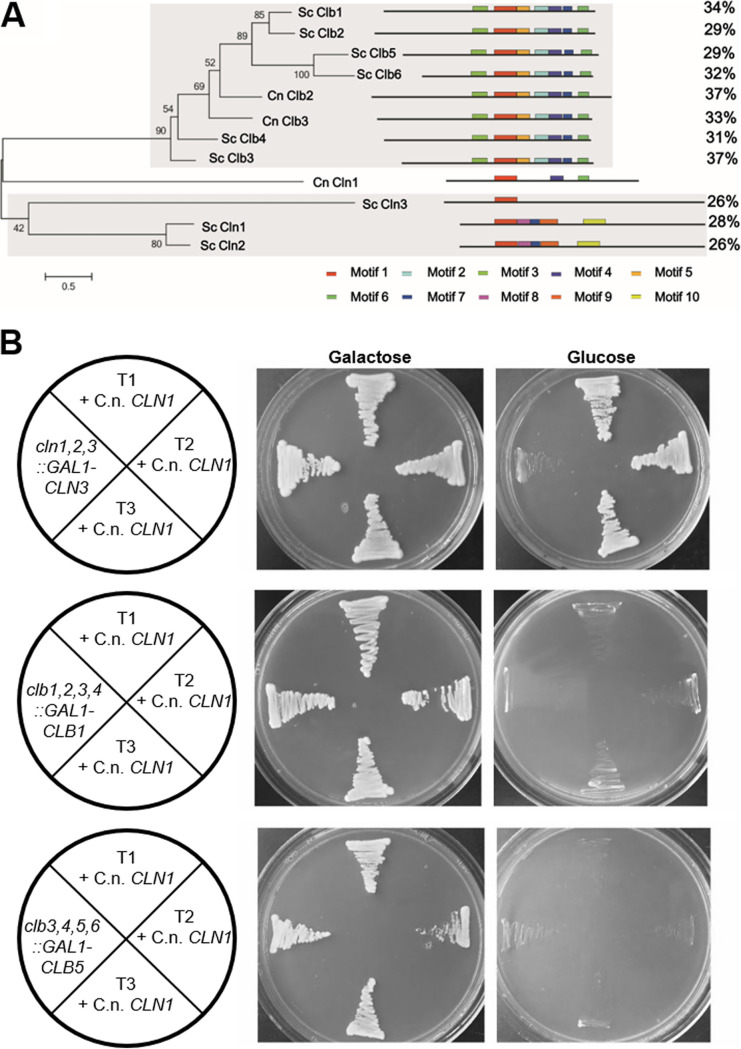
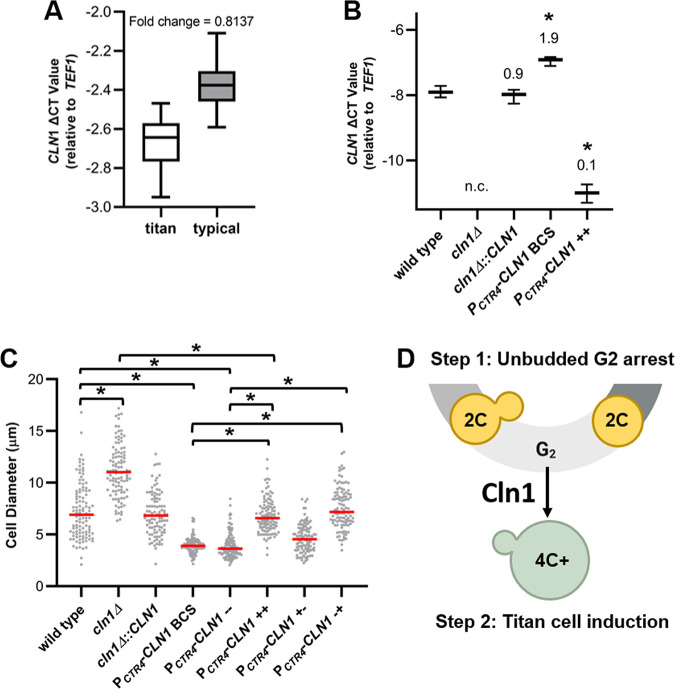
The pathogenic yeast Cryptococcus neoformans produces polyploid titan cells in response to the host lung environment that are critical for host adaptation and subsequent disease. We analyzed the in vivo and in vitro cell cycles to identify key aspects of the C. neoformans cell cycle that are important for the formation of titan cells. We identified unbudded 2C cells, referred to as a G2 arrest, produced both in vivo and in vitro in response to various stresses. Deletion of the nonessential cyclin Cln1 resulted in overproduction of titan cells in vivo and transient morphology defects upon release from stationary phase in vitro. Using a copper-repressible promoter PCTR4-CLN1 strain and a two-step in vitro titan cell formation assay, our in vitro studies revealed Cln1 functions after the G2 arrest. These studies highlight unique cell cycle alterations in C. neoformans that ultimately promote genomic diversity and virulence in this important fungal pathogen. IMPORTANCE Dysregulation of the cell cycle underlies many human genetic diseases and cancers, yet numerous organisms, including microbes, also manipulate the cell cycle to generate both morphologic and genetic diversity as a natural mechanism to enhance their chances for survival. The eukaryotic pathogen Cryptococcus neoformans generates morphologically distinct polyploid titan cells critical for host adaptation and subsequent disease. We analyzed the C. neoformans in vivo and in vitro cell cycles to identify changes required to generate the polyploid titan cells. C. neoformans paused cell cycle progression in response to various environmental stresses after DNA replication and before morphological changes associated with cell division, referred to as a G2 arrest. Release from this G2 arrest was coordinated by the cyclin Cln1. Reduced CLN1 expression after the G2 arrest was associated with polyploid titan cell production. These results demonstrate a mechanism to generate genomic diversity in eukaryotic cells through manipulation of the cell cycle that has broad disease implications.
Keywords: Cryptococcus neoformans; aneuploidy; cell cycle; cryptococcal meningitis; cryptococcosis; cyclins; deneoformans; ploidy; polyploid; polyploidy; titan cell.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
