A Novel Toxoplasma Inner Membrane Complex Suture-Associated Protein Regulates Suture Protein Targeting and Colocalizes with Membrane Trafficking Machinery
- PMID: 34634933
- PMCID: PMC8510555
- DOI: 10.1128/mBio.02455-21
A Novel Toxoplasma Inner Membrane Complex Suture-Associated Protein Regulates Suture Protein Targeting and Colocalizes with Membrane Trafficking Machinery
Abstract
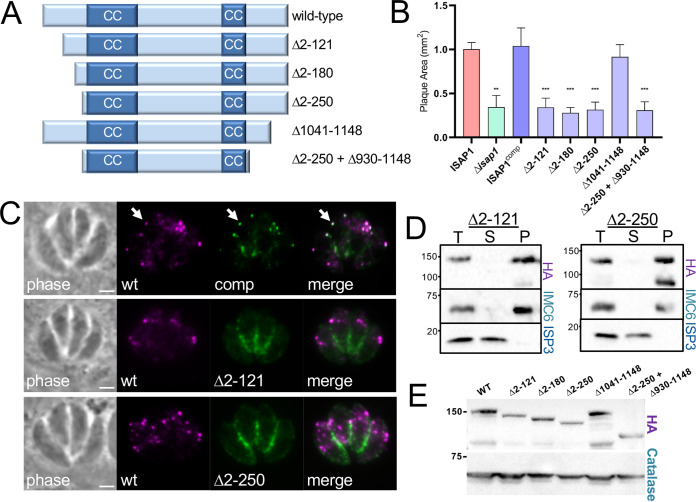
The cytoskeleton of Toxoplasma gondii is composed of the inner membrane complex (IMC) and an array of underlying microtubules that provide support at the periphery of the parasite. Specific subregions of the IMC carry out distinct roles in replication, motility, and host cell invasion. Building on our previous in vivo biotinylation (BioID) experiments of the IMC, we identified here a novel protein that localizes to discrete puncta that are embedded in the parasite's cytoskeleton along the IMC sutures. Gene knockout analysis showed that loss of the protein results in defects in cytoskeletal suture protein targeting, cytoskeletal integrity, parasite morphology, and host cell invasion. We then used deletion analyses to identify a domain in the N terminus of the protein that is critical for both localization and function. Finally, we used the protein as bait for in vivo biotinylation, which identified several other proteins that colocalize in similar spot-like patterns. These putative interactors include several proteins that are implicated in membrane trafficking and are also associated with the cytoskeleton. Together, these data reveal an unexpected link between the IMC sutures and membrane trafficking elements of the parasite and suggest that the suture puncta are likely a portal for trafficking cargo across the IMC. IMPORTANCE The inner membrane complex (IMC) is a peripheral membrane and cytoskeletal system that is organized into intriguing rectangular plates at the periphery of the parasite. The IMC plates are delimited by an array of IMC suture proteins that are tethered to both the membrane and the cytoskeleton and are thought to provide structure to the organelle. Here, we identified a protein that forms discrete puncta that are embedded in the IMC sutures, and we show that it is important for the proper sorting of a group of IMC suture proteins as well as maintaining parasite shape and IMC cytoskeletal integrity. Intriguingly, proximity labeling experiments identified several proteins that are involved in membrane trafficking or endocytosis, suggesting that the IMC puncta provide a gateway for transporting molecules across the structure.
Keywords: Apicomplexa; BioID; Toxoplasma gondii; dynamin-related protein; inner membrane complex.
Figures