

Pannexin-1 and CaV1.1 show reciprocal interaction during excitation-contraction and excitation-transcription coupling in skeletal muscle
- PMID: 34636893
- PMCID: PMC8515650
- DOI: 10.1085/jgp.202012635
Pannexin-1 and CaV1.1 show reciprocal interaction during excitation-contraction and excitation-transcription coupling in skeletal muscle
Abstract
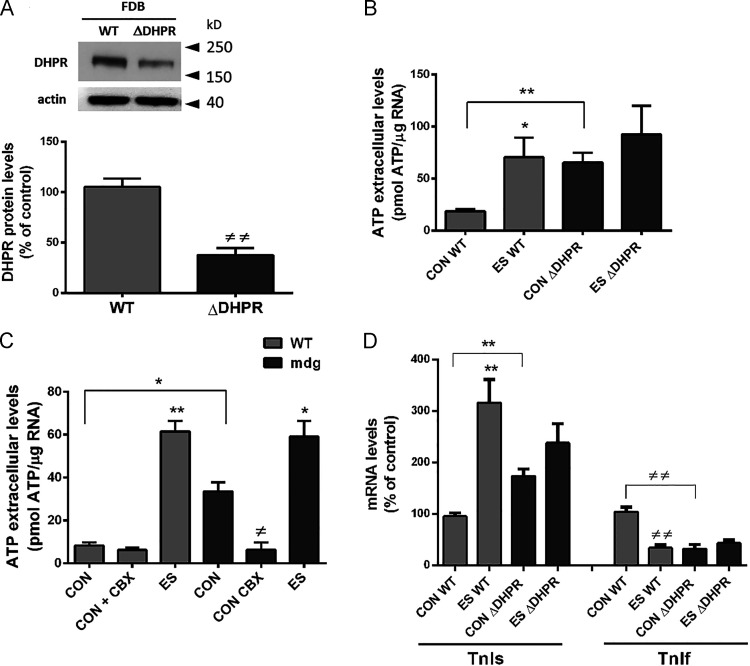
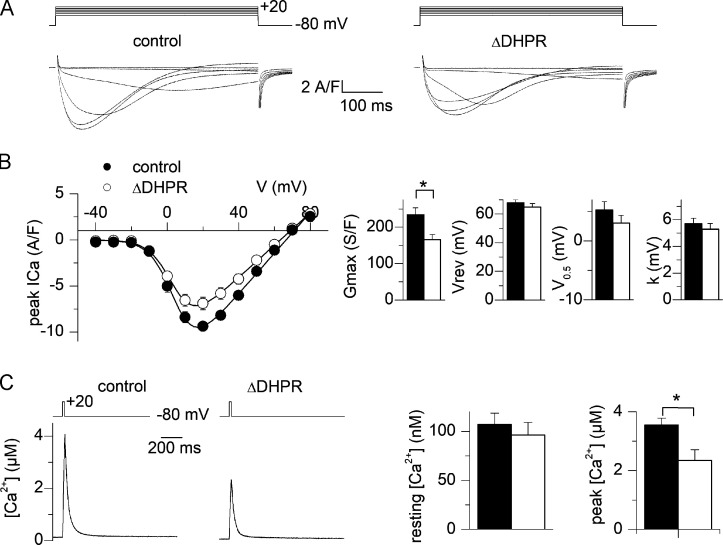
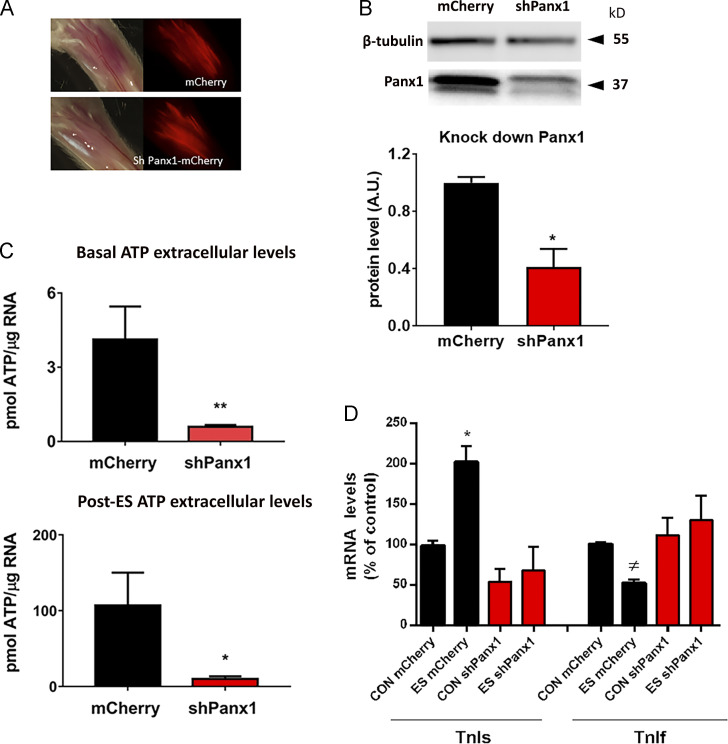
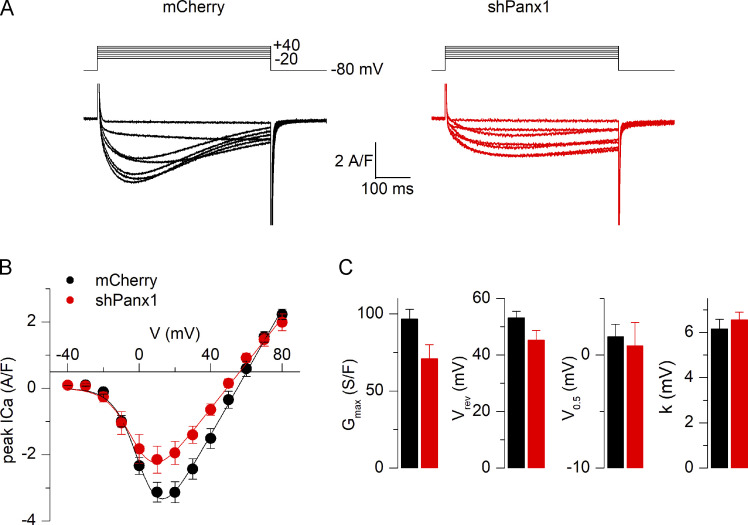
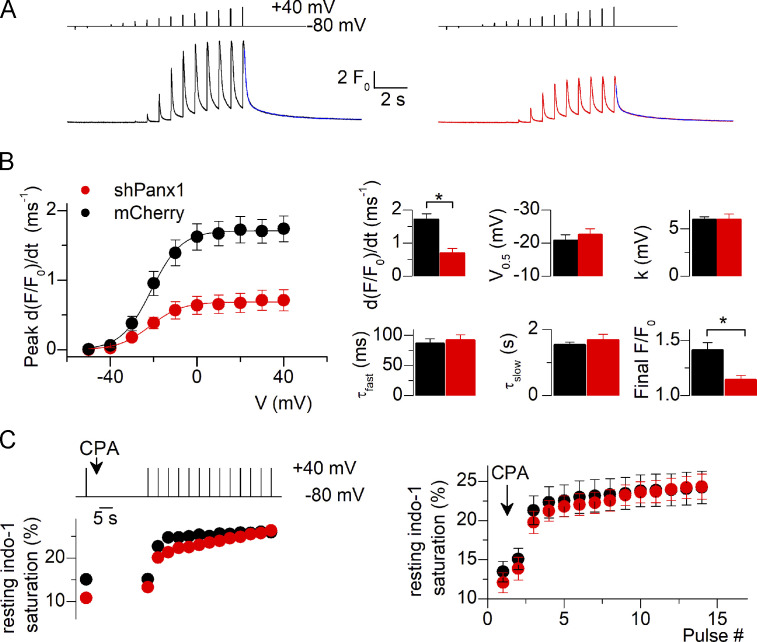
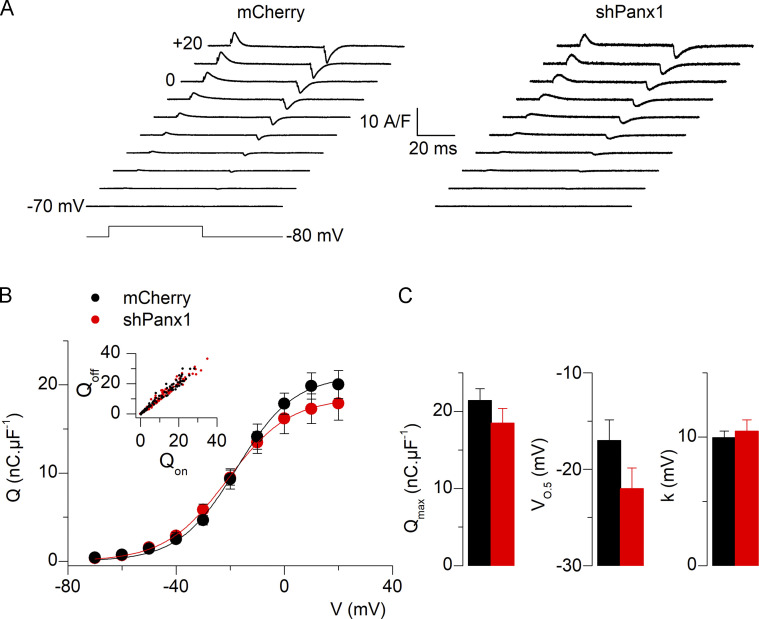
One of the most important functions of skeletal muscle is to respond to nerve stimuli by contracting. This function ensures body movement but also participates in other important physiological roles, like regulation of glucose homeostasis. Muscle activity is closely regulated to adapt to different demands and shows a plasticity that relies on both transcriptional activity and nerve stimuli. These two processes, both dependent on depolarization of the plasma membrane, have so far been regarded as separated and independent processes due to a lack of evidence of common protein partners or molecular mechanisms. In this study, we reveal intimate functional interactions between the process of excitation-induced contraction and the process of excitation-induced transcriptional activity in skeletal muscle. We show that the plasma membrane voltage-sensing protein CaV1.1 and the ATP-releasing channel Pannexin-1 (Panx1) regulate each other in a reciprocal manner, playing roles in both processes. Specifically, knockdown of CaV1.1 produces chronically elevated extracellular ATP concentrations at rest, consistent with disruption of the normal control of Panx1 activity. Conversely, knockdown of Panx1 affects not only activation of transcription but also CaV1.1 function on the control of muscle fiber contraction. Altogether, our results establish the presence of bidirectional functional regulations between the molecular machineries involved in the control of contraction and transcription induced by membrane depolarization of adult muscle fibers. Our results are important for an integrative understanding of skeletal muscle function and may impact our understanding of several neuromuscular diseases.
© 2021 Jaque-Fernández et al.
Figures
References
-
- Al-Qusairi, L., Weiss N., Toussaint A., Berbey C., Messaddeq N., Kretz C., Sanoudou D., Beggs A.H., Allard B., Mandel J.-L., et al. 2009. T-tubule disorganization and defective excitation-contraction coupling in muscle fibers lacking myotubularin lipid phosphatase. Proc. Natl. Acad. Sci. USA. 106:18763–18768. 10.1073/pnas.0900705106 - DOI - PMC - PubMed
-
- Altamirano, F., Valladares D., Henríquez-Olguín C., Casas M., López J.R., Allen P.D., and Jaimovich E.. 2013. Nifedipine treatment reduces resting calcium concentration, oxidative and apoptotic gene expression, and improves muscle function in dystrophic mdx mice. PLoS One. 8:e81222. 10.1371/journal.pone.0081222 - DOI - PMC - PubMed
-
- Arias-Calderón, M., Almarza G., Díaz-Vegas A., Contreras-Ferrat A., Valladares D., Casas M., Toledo H., Jaimovich E., and Buvinic S.. 2016. Characterization of a multiprotein complex involved in excitation-transcription coupling of skeletal muscle. Skelet. Muscle. 6:15. 10.1186/s13395-016-0087-5 - DOI - PMC - PubMed
