

Individual bacteria in structured environments rely on phenotypic resistance to phage
- PMID: 34637438
- PMCID: PMC8509860
- DOI: 10.1371/journal.pbio.3001406
Individual bacteria in structured environments rely on phenotypic resistance to phage
Abstract
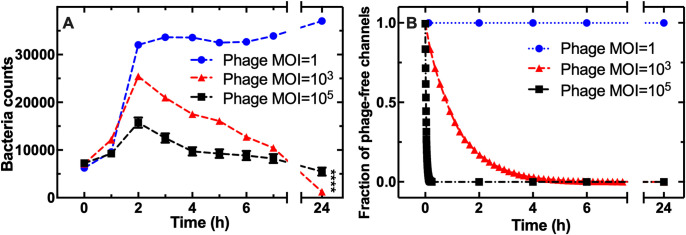
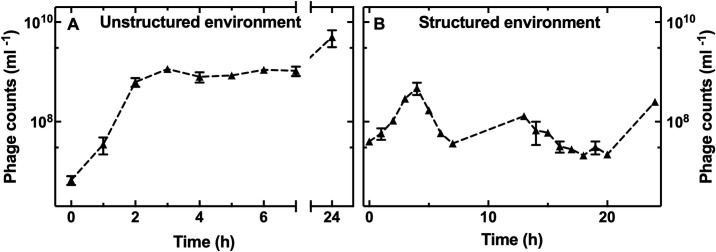
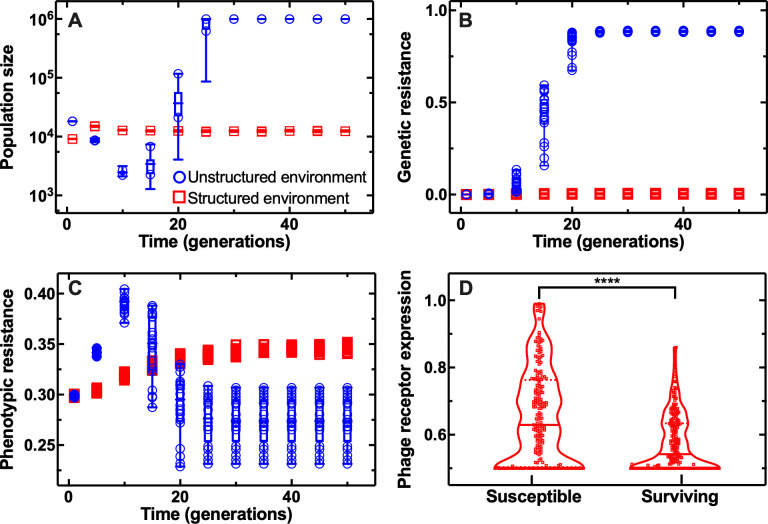
Bacteriophages represent an avenue to overcome the current antibiotic resistance crisis, but evolution of genetic resistance to phages remains a concern. In vitro, bacteria evolve genetic resistance, preventing phage adsorption or degrading phage DNA. In natural environments, evolved resistance is lower possibly because the spatial heterogeneity within biofilms, microcolonies, or wall populations favours phenotypic survival to lytic phages. However, it is also possible that the persistence of genetically sensitive bacteria is due to less efficient phage amplification in natural environments, the existence of refuges where bacteria can hide, and a reduced spread of resistant genotypes. Here, we monitor the interactions between individual planktonic bacteria in isolation in ephemeral refuges and bacteriophage by tracking the survival of individual cells. We find that in these transient spatial refuges, phenotypic resistance due to reduced expression of the phage receptor is a key determinant of bacterial survival. This survival strategy is in contrast with the emergence of genetic resistance in the absence of ephemeral refuges in well-mixed environments. Predictions generated via a mathematical modelling framework to track bacterial response to phages reveal that the presence of spatial refuges leads to fundamentally different population dynamics that should be considered in order to predict and manipulate the evolutionary and ecological dynamics of bacteria-phage interactions in naturally structured environments.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures



References
-
- Pirnay J-P, Verbeken G, Rose T, Jennes S, Zizi M, Huys I, et al.. Introducing yesterday’s phage therapy in today’s medicine. Future Virol. 2012;7:379–90. doi: 10.2217/fvl.12.24 - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
